Эмбриональный, или зародышевый, период развития кеты
В течение зародышевого периода развитие осуществляется за счет запасов, заключенных в желтке, и потому его называют также периодом эндогенного питания. Тот интервал развития, который эмбрионы проводят в икринке, часто называют инкубаци
Существованис дочернего организма начинается со слияния ядер родитсльских гамет и образования зиготы. Этому предшествуют сложные процессы, изученные у кеты достаточно подробно. Осеменение, активация икры, образование зиготы. По данным Ямамото (К. Yamaпoto, 1952), зрелые икринки кеты паходятся на стадии метафазы второго деления созревания. Это типично для зрелых овоцитов костистых рыб (Гинзбург, 1968). Дальнейшее продвижение мейотического процесса осуществляется после встречисперматозонда с яйцом и его активации водой. При температуре 7,5—90° преобразовапие ядра яйцеклетки завершается примерно через 3 час. после осеменения, и женский пронуклеус начипает мигрировать к мужскому, который к этому времени уже достигает места будущей встречи. Через 3,5—4 час. после осемспения пропуклеусы коньюгируют и образуется зигота.
Преобразование ядерного аппарата родительских гамет совпадает повремени с обводнением яиц, образованием перивителлинового пространства и затверденнем оболочки (Зотин, 1954, 1961). Процессоводнения начинается после активации яиц водой и выделения под обволочку осмотически активных веществ, содержащихся в альвеолах кортикального слоя цитоплазмы. Кортикальные альвеолы яицкеты и перки имеют диаметр от 3 до 30 мки содержат осмотически активные кислые мукополисахариды, а также небольшое количество белков и, предположительно, амипосахаров (Kusa, 1949, 1956, 1958. Kanoh, K. Yamomoto, 1957; K. Yamamoto, 1951; T. Yamamoto, 1957; Hannano, 1957).
У лососей полостная жидкость служит мощным активатором и спермии сохраняют в ней подвижность значительно дольше, нсжели в воде (Scheuring, 1925; Doricr, 1951; Гинзбург, 1968). В полостной жидкости и в изотоническом растворс спермии лососей способны проникать в яйца (Привольнев, 1941б). Исследованиями, проведенными на яйцах кеты (Kusa, 1950; K. Yamamoto, 1951) и других лососевых рыб (Гинзбург, 1963, 1968), было показано, что для вовлечения спермия в дальнейшее развитие необходима активация яиц водой. Указанным особенностям половых продуктов отвечает методика «сухого», называемого также русским, способа искусственного оплодотворения икры, разработанного В. П. Врасским. В настоящее время он применяется в несколько измененном виде (Смирнов, 1963б). Икра перемешивается с неразбавленной спермой, которая берется от нескольких самцов. Затем приливается немного воды и перемешивание повторяется. После смены воды, икра выдерживается при полном покое для набухания. В паших опытах при температуре 6—7" и 5"обводнение яиц завершалось в течение от 40—50 до 60 мин. (рис. 6). В результате обводнення весяиц кеты увеличивается на 13,4—18% (Aоki. 1939). Через 10—15 мин. после помещения в водy икринки приобретают слабую клейкость. Они могут соединяться в группы и прикрепляться к частицам грунта, что в какой-то мере предохраLLLLLL G LLL LLLLLLL LLL LLLaLLaS вых ям. При искусственном оплодотворении к икрипкам могут прилипать погибшие сперматоfд 3) 50 0 0" зоиды, сгустки крови, слизь и PHIC, 5, Потребленне BOL LICE MP = другие органические частнцы, KO
неннымен шкрнllкамш кеты торые оказываются субстратом I — прн 6—7" средний вес ик- для развития сапролегнии. КлейS yyLLLLSLSSLSSLS0LS S Ly LkLLL LLLLCktCL LkLLL kLiLiS до набухання 180 мг не воды. Промывку икры следует производить в течение 5—10 мин. после осеменения, пока икра имеет низкую чувствительность к механическим воздействиям. После указанного срока чувствительность икры начинает повышаться (Смирнов, 1955б).
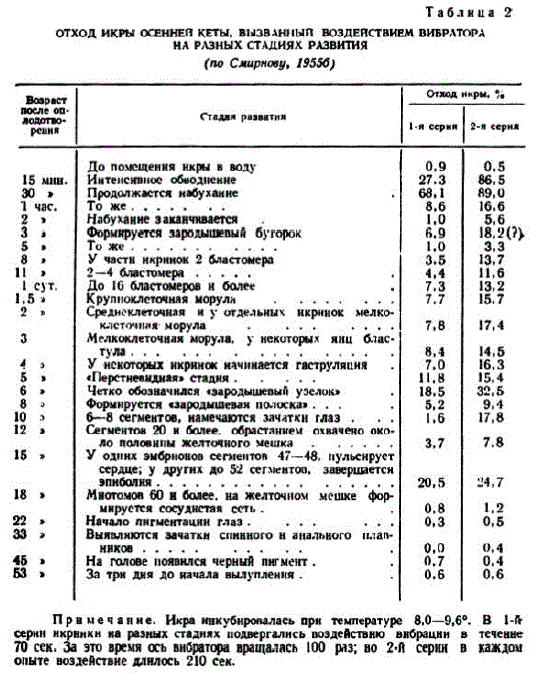
В опытах, данные которых приведены в табл. 2, было показано, что до попадания в поду икра стойко переносит значитсльные механические воздействия. Этим обеспечивастся сохранность икры при толчках самок во время выбивания нcpccтовых ям и закопке икры. Это же качество позволяет успешно транспортировать зрелых наркотизированных или умерщвленных самок, как и неоплодотворенную икру, помещенную в специальную изотермическую тару.
Через 15 мин. после осеменения воздействие вибратора повышало отход, который был особенно велик в опыте, проведенном через 30 мин. Позже чувствительность икры снижалась. Высокая чувствительность совпадала современем обводнения икринок. Одна из очевидных тому причин — ослабление прочности оболочек, другая — ослабление прочности поверхностного слоя цитоплазмы яйцеклетки вследствне разрушения кортикальных альвеол. Набухающая икра должна находиться в полном покое.
У набухших икринок жировые капли сосредоточиваются у анимального полюса, образуя куполообразное выпячивание, на котором формируется цитоплазматический бугорок, или зародышевый диск. Благодаря полярности строения яйцо всегда располагается анимальным полюсом кверху.
Необходимо замстить, что в случаях передержки зрелых самок некоторые икринки могут начать обводняться еще в полости тела, их оболочки затвердевают, жировые капли сливаются, ук
40
Таблнца 2
мируется сосуднстая ссть, O8. l, 2 Начало пнгментацнн глаз. O3 O, 5 33 Выявляются зачатки спинного напального плап
На голове появился черный пигмент. sh, 7 0,4 53 За трн дня доначала вылуплення. 0, 6 O6
Примечание. Икраннкубнровалась прн температуре 8.0—9,6°. В 1-й сернинкрннкн на разных стаднях подвергались воздействню внбрации в теченне T0 сек. это время осъ вибратора вращалась 100 ряз; во 2-й серии в каждом опыте воздействне длнлось 210 сек.
рупняются. Такую перезревшую икру рыбоводы образно называют «горохом». Она не способна оплодотворяться, а ее присутствие указывает и на общее снижение качества икры.
1 этап. Образование цитоплазматического бугорка — зародышевого диска. За начало первого этапа онтогенеза мы принимаем образование зиготы. Продолжается этап до начала дроблепия. На
Тепловском заводе при срcдней температуре 3,4° этот этап длился около 20 час. На Соколовском заводе при 9,6" у одних икринок начало дробления отмечено через 8 час., у других—через 10 час.
TOC Te OCCMCHOHH.A.
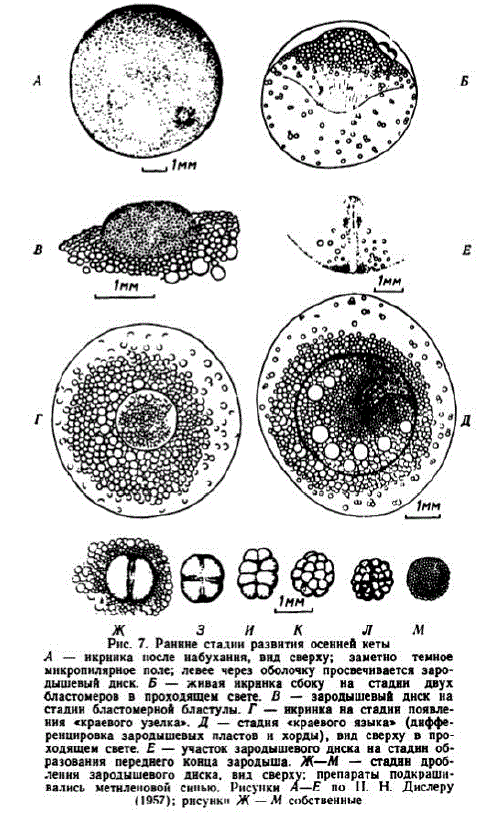
В теченис этапа на анимальном полюсс икринок, в месте скопления жировых капель формируется цитоплазматический бугорок. У зрелых яйцеклеток на анимальном полюсе выявляется лишь нсбольшое скопление цитоплазмы. В псрвые часы после осемснения икринок процссc концентрации цитоплазмы протекаст медленно. В нашем случае через 6 час. после осеменения цитоплазматический бугорок оставался уплощенным и имел диаметр 1,4—1,6 мм, причем контуры его были нечеткими, изломанными, что указывало на незавершенность процесса концентрации цитоплазмы. Сформировавшийся же зародышсвый бугорок принимает форму, близкую к полушарию. Его контур четко очерчивается. Диаметр сформиPಖ್ಖ: зародышевого диска у осенней кеты варьирует от
„9 до 1,3 мм.
К концу этапа нижняя поверхность желточного мешка приобрстает способность прилипать к оболочке (Дислер, 1957). Между поверхностью анимального полюса яйцеклетки и оболочкой образустся пространство, достигающce 0,4—0,8 мм (рис. 7, Б).
В течение этапа чувствительность икры к травмам понижена. Это благоприятствует перевозке набухшей икры с пунктов сбора в инкубаторы и раскладкс ее на рамки. На Сахалине рыбоводы перевозят влажную икру без воды, применяя плотную упаковку. Отход икры за время транспортировки составляет 1—2%.
2 этап. Дробление зародышевого диска. Этап начинается с появления первой борозды дробления и заканчивается образованием бластомерной бластулы, которая формируется при 3,3° в возрасте 8 сут., а при 10,4° в возрасте 3 сут. (около 31 градусо-дня). Образовавшиеся два бластомера по форме напоминают семена бобов. Вторая борозда дробления проходит перпендикулярно первой, образуя четыре плотно прилегающих друг к другу бластомера. Две третьих борозды дробления проходят параллельно первой и образуют 8 бластомеров, располагающихся двумя параллельпыми рядами (рис. 7, Ж—И). Зародышевый диск на этой стадии принимает форму удлипенного овала и его площадь несколько увеличивается. По наблюдению Н. Н. Дислера, у кеты тепловского стада на этой стадии два, три или четыре бластомера лежат пад другими, непосредственно прилегающими к желтку. Борозды четвертого дробления проходят уже в горизонтальной (экваториальной) плоскости и образовавшиеся 16 бластомеров располагаются двумя слоями. Нижние бластомеры крупнее верхних. При просмотре дробящихся нкринок можно видеть не только 4, 8, 16, 32, но также 3, 6, 12, около 30 и иное число бластомеров, что указывает на необязательность синхронного деления всех бластомеров. После 4—5-го дробления зародышевый диск принимает округлую форму.
Тепловском заводе при срcдней температуре 3,4° этот этап длился около 20 час. На Соколовском заводе при 9,6" у одних икринок начало дробления отмечено через 8 час., у других—через 10 час.
TOC Te OCCMCHOHH.A.
В теченис этапа на анимальном полюсс икринок, в месте скопления жировых капель формируется цитоплазматический бугорок. У зрелых яйцеклеток на анимальном полюсе выявляется лишь нсбольшое скопление цитоплазмы. В псрвые часы после осемснения икринок процссc концентрации цитоплазмы протекаст медленно. В нашем случае через 6 час. после осеменения цитоплазматический бугорок оставался уплощенным и имел диаметр 1,4—1,6 мм, причем контуры его были нечеткими, изломанными, что указывало на незавершенность процесса концентрации цитоплазмы. Сформировавшийся же зародышсвый бугорок принимает форму, близкую к полушарию. Его контур четко очерчивается. Диаметр сформиPಖ್ಖ: зародышевого диска у осенней кеты варьирует от
„9 до 1,3 мм.
К концу этапа нижняя поверхность желточного мешка приобрстает способность прилипать к оболочке (Дислер, 1957). Между поверхностью анимального полюса яйцеклетки и оболочкой образустся пространство, достигающce 0,4—0,8 мм (рис. 7, Б).
В течение этапа чувствительность икры к травмам понижена. Это благоприятствует перевозке набухшей икры с пунктов сбора в инкубаторы и раскладкс ее на рамки. На Сахалине рыбоводы перевозят влажную икру без воды, применяя плотную упаковку. Отход икры за время транспортировки составляет 1—2%.
2 этап. Дробление зародышевого диска. Этап начинается с появления первой борозды дробления и заканчивается образованием бластомерной бластулы, которая формируется при 3,3° в возрасте 8 сут., а при 10,4° в возрасте 3 сут. (около 31 градусо-дня). Образовавшиеся два бластомера по форме напоминают семена бобов. Вторая борозда дробления проходит перпендикулярно первой, образуя четыре плотно прилегающих друг к другу бластомера. Две третьих борозды дробления проходят параллельно первой и образуют 8 бластомеров, располагающихся двумя параллельпыми рядами (рис. 7, Ж—И). Зародышевый диск на этой стадии принимает форму удлипенного овала и его площадь несколько увеличивается. По наблюдению Н. Н. Дислера, у кеты тепловского стада на этой стадии два, три или четыре бластомера лежат пад другими, непосредственно прилегающими к желтку. Борозды четвертого дробления проходят уже в горизонтальной (экваториальной) плоскости и образовавшиеся 16 бластомеров располагаются двумя слоями. Нижние бластомеры крупнее верхних. При просмотре дробящихся нкринок можно видеть не только 4, 8, 16, 32, но также 3, 6, 12, около 30 и иное число бластомеров, что указывает на необязательность синхронного деления всех бластомеров. После 4—5-го дробления зародышевый диск принимает округлую форму.
42
В процессе дальнейшего дробления верхние бластомеры располагаются плотнее, они немного мсльчc и уплощены. Нижние бластомеры крупнее, делятся медленнее, расположены рыхло. Те из них, которые лежат на периферии бластодиска, сохраняют связь с краевой цитоплазмой. Часть цитоплазмы нижних бластомеров заходит между глыбками желтка, давая начало центральному участку перибласта.
Под зародышсвым диском и в непосредственной близости от него расположены более мелкис жировые капли. Если яйцо быстро перевернуть, они перемещаются вместе с бластодиском. Болсс крупные жировые капли, расположснные понсриферии, перемещаются в желточном мешке легко, независимо отбластодиска (Дислер, 1957).
С началом дробления повышается чувствительность икры к механическим воздействиям (см. табл. 2). Поэтому к началу дробления икра должна быть размещена в аппараты и инкубироваться при полном покое.
3 этап. Бластула. Продолжается от образования бластомерпой бластулы до начала гаструляции. При температуре 3,3" этап длится 10—11 сут., при 10,4° — 2,5—3 сут. Одним из внешних признаков перехода к стадии бластулы служит начало увеличения бластодиска (Иванов, 1937; Дислер, 1957). У ксты диаметр бластодиска в это время составляет 1,3—1,4 мм. Он куполообразно выдается над поверхностью желточного мешка и охватывает зону мслких жировых капель.
На стадии бластулы хорошо выражен псрибласт, формирующийся в результате деления бластомеров, прилегающих к жслтку, При этом одно из повых ядер образует типичный бластомер, а второс ядро проникаст в промсжуточный слой цитоплазмы, отделяющсй бластодиск от желтка. При дальнейшем митотическом делении этих ядер подбластодиском образуется многоядерное синцитиальнос образование, пазываемос перибластом, или параблacTo M (Riddle, 1917; PMB a HoB, 1937; Manon, Hoar, 1956).
Поверхность бластулы покрыта слоем плотно прилегающих друг к другу клеток полигональной формы, под которыми рыхло располагаются более крупные клетки. Начальную стадию бластулы благородноголосося П. П. Иванов назвал бластомерной бластулой. В дальнейшем наружный слой клеток уплотняется и к нему прилегают нижележащие бластомеры, наблюдается процесс эпителизации. Поздняя бластула названа эпителиальной бластулой. Диаметр поздней бластулы у осенней амурской кеты достигает 2,3—2,6 мм (Дислер, 1957). Яйца сахалинской осенней кеты крупнее, и у нее диаметр бластулы достигает 3,0—3,2 мм.
4 этап. Образование зародышевых пластов. В течение этапа протекает процесс гаструляции. В условиях Соколовского завода при средней температуре 10,4° образование зародышевых пластов начиналось на исходе шестых суток. Этап закончился через двое суток, когда образовались первые мезодермалыIые сегменты. На
44Тепловском заводе при 3,4° гаструляция начинается в возрасте 18—19 сут. и завершается примерно черcз 6 сут.
Во время гаструляции зародышевый диск увеличивастся, уплощается, в центральной части становится тонким. В одпом из краевых участков бластодиска, будущем каудальном конце формирующегося зародыша, образуется утолщение — «краевой узелок» (рис. 7, Г). Методом прижизненной маркировки участков бластодисков установлено, что поверхностные клетки бластодермы у лососевых и других костистых рыб распространяются по поверхпости желточного мсшка исключитсльно путем эпиболии, не подворачиваются под зародышевый диск и не принимают участия в формировании гипобласта. Процесса инвагинации, описывавшегося ранее (Pasteels, 1936), у костистых рыб не наблюдается. В определенное время внутренние клетки разрастающейся бластодермы поодиночке или группами отделяются от нес, мигрируют в направлении зачатка тела и образуют гипобласт, а клетки, лежащие непосредственно под эпителиальной поверхностью бластодермы, смещаясь единым пластом, формируют эпибласт (Ballard, 1966, 1968; Ballard, Dodcs, 1968; TpHHKayc, 1972).
При диаметре зародышевого диска около 3,5 мм его края псреходят зону расположения мелких жировых капель. «Краевой узслок» удлиняется и превращается в «зародышевый язычок». Зародышевые пласты обособляются четчc. По данным Манон и Хора (1956), ့ဖူးဗုဝှိုးဂံ слой зародыша представлен уплощенными клетками. Подним лежат более крупные клетки эктодермы. Непосредственно под эктодермой, по продольной оси зародыша, плотным тяжем располагаются клетки мезодермы. Между ними и перибластом разбросаны клетки энтодермы, которые несколько позже уплощаются. В переднем конце зародыша эктодерму подстилает выделяющийся слой клеток прехордальной пластинки. В ядрах центрального перибласта митоза ужс не наблюдается, они делятся амитотически. В начале гаструляции Манон и Хор обнаруживали группу заметно увеличенных клеток с выростами, подобными псевдоподиям амебы. Эти амсбообразные клстки предположительно были названы примордиальными зародышевыми клетками. Г. М. Персов (1966, 1968) относит появление первичных половых клеток у кеты к песколько более позднему возрасту.
Когда диаметр зародышевого диска достигает 4 мм, зародышевая полоска увеличивается до 1,5 мм. На поперечных срезах такого зародыша, сделанных несколько впереди его каудального выступа, в центральной части выявляются концентрически располагающиеся клетки зачатка хорды. По бокам от него лежит недифференцированная боковая мсзодерма. Под зачатком хорды инсмного сбоку образуется полость, окруженная цилиндрическими клетками, представляющая зачаток Купферова пузырька. Снаружи по сторонам продольной оси эмбриопа утолщенный слой эктодермальных клеток образует небольшнс продольные выпячивания — нервную пластинку..
Зародышам рыб на ранних этапах развития свойственна протоплазматическая моторика, наблюдавшаяся многими исследователями. У благородноголосося эмбриональная моторика исследовалась методом цейтраферной киносъемки (Willker, 1953). Изучены закопомерности изменений эмбриональной моторики в онтогенcзерыб разных экологических групп и ее зависимость от температуры и газового режима (Рcзниченко, 1959а, б; Резниченко и др., 1968). Движения яиц под оболочкой обусловлены сократительной активностью цитоплазмы, покрывающей желточный мешок. Волны ес сокращений вызывают деформацию и смещения центратяжести яйца, что приводит к изменению его положения в пространстве. У лососей на ранних этапах развития анимальный полюс яйца описывает круговое движение по направлению или против движения часовой стрелки. При низкой температуре уловить эти движения, наблюдая живое яйцо под микроскопом, трудно. На данной стадии движения становятся заметнсе. Во время наших наблюдений при 14°бластодиск ксты описывал полный «круг» вращения за 4 мин. 40сек. — 5 мин. В результате подобных движений яйца, а позже и самого эмбриона перемешивается перевителлиновая жидкость и интенсифицируется внешнее дыхание зародышей.
С началом гаструляции чувствительность икры к механическим воздействиям возрастает и одинаковое воздействие на стадии хорошо выраженного «зародышевого узелка» вызывает примерно вдвое больший отход, чем послс воздсйствия на стадиях дроб
(IIHH.
5 этап. Формирование головы и туловища зародыша. Этап охватывает интервал развития от начала сегментации туловищной мезодcрмы до образования 46—47 сегментов. При средней температуре 32 он начиналcя на 24-25-e сут и заканчивалcя на 35—36-е сут. В условиях Соколовского завода начало сегментации отмечсно на исходе 8-х — вначале 9-х сут. (81—85 градусо-дней). К исходу 10-х сут. зародыши имели до 18—20 сегментов, в 12 сут. — 35—36 сегментов. Этап закончился в возрасте 14 сут. (137— 142 градусо-дня).
Пcрвые сомиты формируются у зародышей длиной 2 мм, когда диаметр зародышевого диска увеличивается до 4,5 мм (Мапоп, Ноаг, 1956). Н. Н. Дислер образование трех сомитов и начало дифференцировки отделов головного мозга отметил при длине эмбриона 2,5 мм. С достижением длины 3 мм число сомитов возрастало до семи (при 3,4° на 26-е сут.). Бластодиск к этому времени охватывал до половины окружности яйца. У эмбрионов длиной около 3,5 мм число сегментов возрастало до 13 (рис. 8, А—В).
Манон и Хор указывают, что при длине эмбриопа 3,0—3,5 мм в зачатках глазобразуются полости, они превращаются в глазные пузыри. Вскоре дистальная часть их стенки инвагинирует и образуются глазные бокалы. Над передним концом хорды благодаря инвагинации внутренних слоев эктодермы появляются зачатки слу
ховых пузырьков. В момент закладки они удалены от переднего конца зародыша на значительное расстоянис. В основании головного мозга на уровне заднего края глаз закладывается воронка (infuпdibuluпn). Несколько позади этой области эктодермальный зачаток подгибается внутрь, давая начало глоточным карманам. Из пижнего участка мезодермы начинает формироваться зачаток сердца. Mcзодермальные боковые пластинки делятся на соматический и сплапхнический слон, узкая щель мсжду которыми указывает на образование перикардиального целома. Продольное скопление энтодермальных клеток в средней части тела зародыша обpa3yeT 3avHaToK KHLLIKH.
У зародышей длиной 4 мм сомиты дифференцируются надерматом, миотом и склеротом. Формируется пронефрическая складка. Вентролатеральнее склеротома вдоль тела располагаются клетки, дающие начало осевым кровеносным сосудами клеткам крови.
На Тепловском заводе на 31-е сутки развития, когда зародыши достигли длины около 4,0—4,5 мм, у них насчитывалось 30—31 сомит и бластодермой покрылось более */, желточного мсштка. При витальном микроскопировании были хорошо видны дифференцировка отдслов головного мозга и энцефаломеры продолговатого мозга, глазныс бокалы, плакоды хрусталиков, слуховые пузырьки. Зачаток сердца в виде трубки опустился в обширную перикардиальную полость (рис. 8, Г).
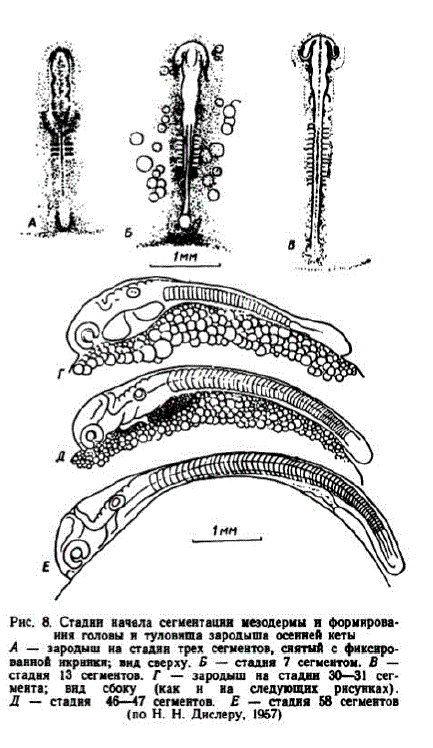
По данным наших опытов, с того времени, когда бластодермой покрывается болсс половины поверхности желточного мешка, чувствительность икры к толчкампа некоторое время снижается. 6 этап. Обособление задней части тела зародыша от поверхности желточного мешка. По данным Н. Н. Дислера, этап продолжается от формирования 46—47 до 65 сегментов. На Тепловском заводе при средней температуре 3" этап длился с 35—36 до 45 сут. В течение этапа длина зародышей увеличивалась от 4,8 до 6,5— 7.0 мм. На Соколовском заводе при средней температурс 9,6°этап пачинался в 15-суточном возрасте, а при 10,1° — на исходс 14 сут. и длился 4—4,5 сут. Существенная функциональная особенность этапа состоит в становлении нервномышечной моторики эмбриоHOB.
Внешний вид зародыша в начале этапа показан на рис. 8, Д. К этому времени процесс эпиболии — обрастание желточного мсшка бластодермой — приближался к завершению. Начала дифференцироваться хвостовая почка. Впереди глаз выявились обонятельные плакоды. Намстились зачатки жаберных крышек. Сердечная трубка становится изогнутой, наблюдаются редкие, слабые сокращения ее стенок. Частота пульсаций сердца быстропарастает, обеспечивая перемешивание полостной жидкости. Миотомы приобрели способность сокращаться. Потребленис кислорода икрой заметно увеличнвается (см. табл. 3), что отвечает возрос
шим потребностям эмбрионов, которые достигли значительного размера.
47

нрованне головын туловнцця 4.0 Обособленне задней части тела. (), 76 4.0 Начало кровообращення . . . . . . 1 Աl 40 Подкишечно-желточное кровообращение 2,88 40 Печеночно-желточное кровообращение. 3,39 4. Перед вылуплением , 4. 45 Во время вылупления . . . . . . . . 7,34 45-50 Более позднне вылупившнеся зародыши 9,5 5,0-60 Jlrq HHKH 32 BO-90 Мальки 33.5
"Прн меч анне. Ошыты проводились с 3-кратной повторностью при содертанникнслорода: до вылуплення б—8 мг/л, у спободных зародышейнлнчинок — 7-8 мг/л, у малыков — прн 8—10 мг/л.
Мелкие жировые капли располагаются преимущественно под головной областью зародыша. Они и небольшое количество прилежащих к зародышу крупных жировых капель при повороте икринки перемещаются вместе с зародышем. Жировые капли, отстоящис дальшс от тела эмбриона, при поворотс икринки меняют свое положение в желтке независимо от его расположення.
У зародыша длиной 5 мм зачатки хрусталика еще не отделсны от эктодермы. В глазах начинают дифференцироваться ретинальный и пигментный слои. Обозначилась закладка мозжечка. Видны три пары глоточных карманов. Формируется эпимиокард.
Заложились осевые кровеносные сосуды и в перикардиальной полости появились эмбриональные кровяные клетки. Вдоль средней линии тела выявляются зачатки органов боковой линии.
После отчленения хвостовой почки быстро нарастает число хвостовых сегментов. Хвост удлиняется, на нем появляется зачаток непарной плавниковой складки. С началом отделения от желтка задней части туловищного отдела развивается преанальная CKT ALKA,
На Тепловском заводе в возрасте 42 сут. (147 градусо-дней) у зародышей насчитывалось до 58 сегментов, из которых 16—17 хвостовых. Длина зародышей достигала 5,5—6,0 мм. У них сформировался задпий отдел кишки и образовалось анальнос отверстие (рис. 8, E). Аналогичного состояния на Соколовском заводе зародыши достигали в возрасте 15 сут. (150 градусо-дней), имея длину 6,0—6,3 мм. Процесс эпиболии в условиях Тепловского завода завершался на 45 сут. (156 градусо-дней), а Соколовского — на 18-с сут. (183 градусо-дня). Во время закрытия желточной пробки икринки кеты и других лососей чрезвычайно чувствительны к толчкам и погибают даже от слабых прикосновений. Эпиболия завершается у разных зародышсй не одновременно и пока этот процесс не закончится, икра должна находиться в абсолютном TOKOHE.
Эмбрионы кеты имеют очень большой желточный мешок и его обрастание затягивается до поздних стадий развития. В самых мелких икринках кеты эпиболия завершается при 52— 53 сегментах, тогда как у зародышей S. salar при наличии около 30 cer MehToB (Battle, 1944).
После завершения процесса эпиболии чувствительность икры к травмам резко падает. Даже у кеты это происходит раньше появления пигмента в глазах, что полезно учитывать рыбоводам. К моменту исчезновения желточной пробки хрусталики отделяются от эктодермы и погружаются в глубь глазных бокалов. Значительно продвигается дифференцировка головного мозга: увеличиваются его желудочки, закладываются обонятельные доли, формируются крупный мозжечок, эпифиз и гипофиз. Намечается дифференцировка первых жаберных дуг. Энтодерма образует кишечную трубку. Пальцеобразный вырост средней кишки дает начало печени. Начинают формироваться почечные клубочки и пронефрический проток. На желточном мешке появляются кропяIIыс островки.
К концу этапа задняя часть туловищного отдела до 29-го сегмента отделяется от желточного мешка. В течение этапа увеличивастся подвижность эмбриопов. Они поводят из стороны в сторону хвостом и временами совершают им волнообразные дви
HKCIIHH.,
7 этап. Развитие подкишечно-желточной системы кровообращения. Этап охватывает интервал от начала кровообращения до формирования кардинальных вен. При средней температуре 9,6°
он длится 1,5—2 сут., при 3,5°— около 5 сут. К концу этапа эмбрионы достигают длины 7,3—7,8 мм и у них формируется до 71 сегмента.
В условиях Соколовского завода начало кровообращения у некоторых эмбрионов отмечалось при 52—53 сегментах, когда еще имелась небольшая желточная пробка (15 сут., 144 градусодня). У основной массы эмбрионов это происходило на 16— 17-е сутки, когда число сегментов возрастало до 58—60. В начале кровообращения ток крови в сосудах слабый, эритроцитов мало и они имеют светлую окраску. Гистохимически присутствие гемоглобина у лососей выявляется до начала тока крови (Остроумова, 1962).
В начале этапа от сердца кровь тсчет по парным мандибулярным дугам аорты — агс. папdibularis aогtae. Над жаберной областью от них ответвляются сосуды, по которым кровь течет в голову и, по спинпой аорте — aогta doгsalis в туловище. Позади ануса спинная аорта переходит в хвостовую артерию — а. caudalis. В начале кровообращения хвостовая артерия проходит лишь вдоль 2—3 хвостовых сегментов, изгибается книзу и переходит в хвостовую вену — v. caudalis. Этот сосуд, обогнув заднюю кишку, непосредственно переходит в подкишечную вену — v. subintestiпаlis, по которой кровь течет на протяжении 25—26 сегментов. Чуть впереди преанальной плавниковой складки подкишечная вена выходит на левую сторону поверхности желточного мешка, давая начало желточной вене — v. vitellinae, которую называют также подкишечножелточной веной — v. subintestinalis vitellinae (Соин, 1968). На левой стороне желточного мешка поток крови описывает небольшую дужку, объединяется с кровью, идущей из передней кардиналышой вены — v. сагdinalis апtегіог и попадает в предсердие. У других лососей первичный круг кровообращения образуется аналогично.
В условиях Тепловского завода у кеты пачало кровообращения отмечалось в возрасте 45 сут., тотчас после закрытия желточной пробки. Эмбрионы имели длину от 6,5 до 7,2 мм, у них насчитывалось 65 сегментов, из которых 41 в туловище. К этому времсни закладывались грудные плавнички. Голова начинала отделяться от поверхности желточного мешка. Уже в начале этапа кровь текла по хвостовой вене до 10-го хвостового сегмента и у заднего участка подкишечно-желточной вены имелись два или три капиллярных сосуда (рис. 9, А, Б). Это более поздняя стадия дифференцировки. Задержку начала кровообращения можно объяснить меньшим размером икринок и лучшей аэрацией воды, подаваемой в инкубатор Тепловского завода.
В течение этапа хвостовые артерия и вена удлиняются, подкишечно-желточная вена опускается ниже, число капилляров на Желточном мешке увеличивается. Жcлточная вена смещается ВНИз следующим образом. Постепенно количество форменных Элементов в кровяных островках, расположенных вблизи внеш
ней стороны вены, увеличивается, они сливаются и включаются в кроветок, тогда как внутрений участок сосуда постепенно запустсвает. Новые капилляры образуются также за счет слияния кровяных островков (Дислер, 1957).
Развивающиеся зародыши двигаются все энергичнее и чаще. С началом кровообращения потребление икрой кислорода по сравнению с предшествующим этапом почти удваивается. Икра стойко переносит механические воздействия (см. табл. 2).
8 этап. Возникновение кардинальных вен и смешанного подкишечно-желточного и печеночно-желточного кровообращения. При развитии в инкубаторе Тепловского завода существенные измснения эмбрионов отмечены на 50-й день развития (около 160 градусо-дней), а на Соколовском заводе — в 18—19-суточном возрасте (183—193 градусо-дня). В первом случае этап длился около 14 сут., во втором — 5 сут. К его завершению эмбрионы достигали 10,5—11,0 мм длины.
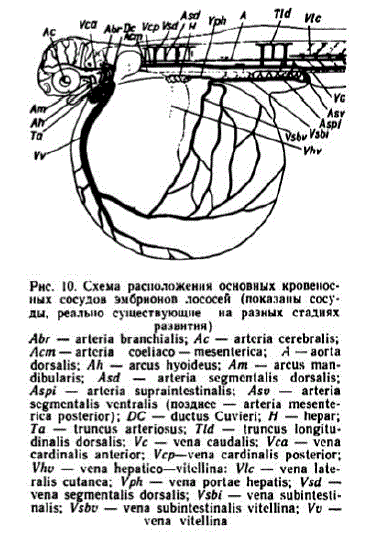
К началу этапа сосуды обильно заполняются эритроцитами, имеющими интенсивную окраску. Начинается кровоснабжение глаз. Из головы кровь отводится передними кардинальными венами — v. cardinalis anterior, которые впереди грудных плавников сливаются с возникшими задними кардинальными венами — v. cardinalis posteriог и образуют короткие кювьеровы протоки — d. d. Cuvieri (рис. 10). В сосудах двух первых пар жаберных дуг появляются кровяные элементы, потока крови в них еще нет. Назначительной части задней половины желточного мешка образуется сеть капиллярных сосудов. Они имеются теперь не только на левой, как было ранее, но и на правой стороне. У живых зародышей позади грудных плавничков хорошо просматриваются клубочки пронефроса.
На Тепловском заводе через 3—5 сут. после начала этапа, а в нашем случае через 1—1,5 сут. сегментация хвоста заканчивается и число миотомов достигает максимума — 72—73, из которых 30—31 хвостовых. Длина эмбрионов — 8—8,2 мм. К этому времени заметно вырастают грудные плавники. На спине непарная плавниковая складка разрастается до первых миотомов. Голова обособляется от желточного мешка. В связи с образованием изгибов головного мозга слуховые пузырьки сблизились с глазами и длина головы yмeньшилась. В слуховых пузырьках идет формирование полукружных каналов, образовалось по нескольку мелких отолитиков. В глазах можно заметить зерна меланина. Формируются обонятельные ямки. Появился ток крови в первых парах жаберных дут. На желточном мешке кровеносные сосуды заняли значительную площадь. Краевая желточная вена опустилась на нижнюю сторону желточного мешка. Печень достнгла довольно крупного размера, расположена между 5 и 9 сегментами, причем сдвинута немного вправо. Возникло печеночно-желточное кровообращение. Первоначально из печени на левую сторону желточного мешка кровь течет по одному тонкому сосуду
и впадает в краевую желточную вену. На правую сторону желточного мешка из печени выходят два сосуда. Первый образует короткую петлю и впадает в правый кювьеров проток, кровь из второго сосуда течет к краевой желточной вене. Эти сосуды вскоре начинают ветвиться (рис. 9, В, Г). Подкишечно-желточная
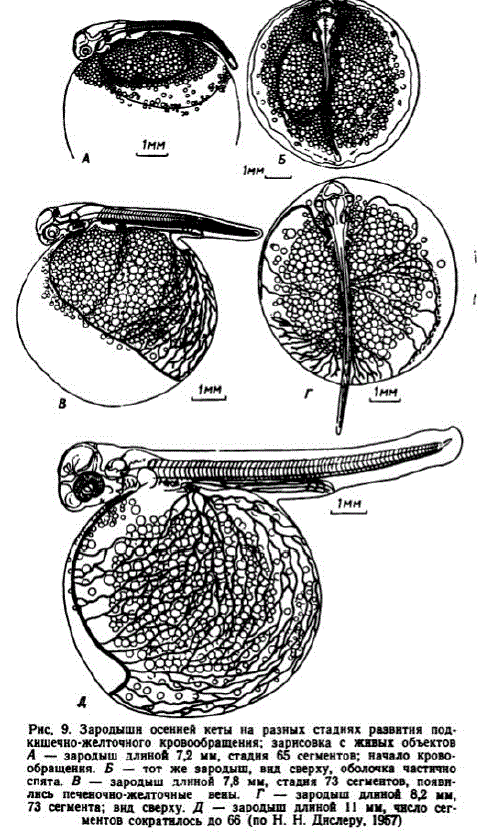
Pнс. 10. Схема расположения основных кровеносных сосудов эмбрионов лососей (показаны сосуды, реально существующие на разных стадиях развитин) Abr. = arteria branchialis; Ac = arteria cerebralis; McPI = arteria coeliaco = mesenterica; A = aorta dorsalis; M h - arcus hyoideus; Am -- arcus mandibularis; Asid - arteria seg Tn Cintalis dorsalis; Aspi - arteria Supra intestinalis: ASU - arteria scg ncintalis vcntralis (n03 AHCC - arteria mesenterica posterior); DC - ductus Cuvieri; H - hepar; Ta - truncus arteriosus; Tld - truncus longitudinalis dorsalis; W - Wena caudalis; Wra - Wena Cardinalis an terior; Warp-wena Cardinalis posterior; WhU - Wena ಙ್ಗico್ನಗ್ಧ W H Wen a lateralis cutanca: Vph — viena portae hepatis; Vsd -- Vena Segmentali5 dorSalis; Vsbi - vena subintestinalis; Visby - wena Subin testinalis witcellina; Wu - vепa Vitellina
вена, снабжающая кровью сосуды каудального участка желточного мешка, после образования задних кардинальных вен и печеночного кровообращения постепенно запустевает. Вместе с тем на желточном мешке возрастает количество сосудов, снабжаемых кровью из печени. Таким образом, для описываемого этапа характерно смешанное кровоснабжение желточного мешка при постепенном возрастании роли печеночного кровоснабжения.
У кеты, развивающейся в инкубаторе Соколовского завода, в возрасте 21 суток (205 градусо-дней) в цитоплазме некоторых
эпителиальных клеток в области формирующихся ротовой и жаберной полостей обнаружены гранулы секрета полисахаридной природы. Предполагастся, что эти клетки выделяют в подоболочечное пространство фермент гиалуронидазу (Сакун, Чистова, 1960). Этот фермент обнаружен в перивителлиновой полости икринок кеты. Его функцию связывают с рсгулированием проницаемости яйцевых оболочек и покровов эмбрионов, а также с участием в акте вылупления (Бузников, Игнатьева, 1958).
9 этап. Функционирование печеночно-желточной системы кровообращения. Этап длится с переключения функции кровоснабLHGaL HLLL LL LLLLL LLLL SLLLLLLLL L S aLLLLLLL LLLL LL LS чала дифференцировки конусов миотомов. На Тепловском заводе он начинался на 64-е сут. (224 градусо-дня) и продолжался около 19 сут., на Соколовском — в возрасте 23—24 сут.
(235—245 градусо-дней) и заканчивался через 6—7 сут.
К началу этапа увеличивается высота головы зародышей (рис. 9,Д). Слуховые капсулы приближаются к глазам. Глаза становятся темными. Многочисленные мслкие отолитики объединяются в крупные образования. Намстившаяся ранес воронка ротовой полости углубилась. Ускорился рост нижней челюсти. Заметно разрастаются жаберные крышки. Основания грудных плавников образуют с продольной осью тела угол около 30°. Удлинившийся хвостовой отдел превышаст 1/3 общей длины эмбриона. Задний конец хорды изогнулся вверх и хвостовой плавник утратил симметричность. Начали формироваться опорные и мышечные элементы хвоста (рис. 11,A). Число хвостовых сегментов сокращается до 27—28. Аналогичный процесс свойствен и другим костистым рыбам. В хвостовом отделе идет обособление позвопков. Содержимое кишечника приобрело лимонно-желтую окраску, обусловленную, по-видимому, появившимися желчными пигменTMP
У эмбрионов четко различаются предсердие и желудочск. Обнаруживается движение крови в гиоидных дугах, в третьей, а немного позже и в четвертой парах жаберпых дуг. Развивается сеть капилляров в покровах теменной области головы, затем над мозжечком и в затылочной области. Появляются сегментальные сосуды, формирование которых идет от головы к хвосту, Хвостовая вена утрачивает связь с подкишечной, а последняя — с сосудистой системой желточного мешка. Задний отдел кишки оплетают разветвления надкишечной артерии (а. supraintestinalis), а средний отдел — капилляры кишечной артерии (а. сосliaca). Из этих капилляров кровь поступаст в подкишечную вену, по которой направляется к печени. На желточном мешке развивается мощная сосудистая сеть, снабжаемая кровью исключительно через печеночные вены.
При длине эмбрионов около 13 мм в месте закладки анального и спинного плавников обнаруживается скопление мезенхимы и зачатки мускульных почек. Удлинившийся хвостовой отдел до
стигает 37% общей длины эмбриопа. В месте закладки опорных LLLLLLL L0LLLLLLL0 LGGLLLLLLLLCLLLLLLL0 LLLLCLL GG S G0CLLCCLGHLHLCLS LLLLCLLL капилляров. Капилляры покрывают всю поверхность желточного мешка. У извлеченных из икринок зародышей желточный мешок принимает яйцевидную форму. Диаметр некоторых жировых капель достигает 1 мм.
10 этап. Дифференцировка верхних и иижних конусов миотомов. Верхние и нижние конусы миотомов при средней температуре 3,3° начинали формироваться на 83-й день (около 270 градусодней). В нашей серии это происходило на 30—31-е сут. (207— 217 P೫೦:೫ಣ್ಣಿ При температуре 3,3" этап длился 12—17 сут., а при 10° - 5-6 сут.
В начале этапа эмбрионы имеют длину 14 мм и более (рис. 11, Б). К этому времени жаберные крышки стали прикрывать 2-ю пару жабер. На 3-й, а затем 4-й и первой парах жаберных дужек появляются зачатки жаберных лепестков. Основания грудных плавников принимают почти вертикальное положение и они приобретают подвижность. Под 25—30 туловищными миотомами появляются небольшис продольные складочки зачатков брюшных плавников. В основания закладывающихся непарных плавников врастают выступы конусов миотомов, образующие впоследствии мышцы плавниковых лучей. Зачатки лучей первоначально представлены стержневидными сгущениями мезенхиматозных клеток. Процесс охрящевения приводит к образованию лепидотрихий, которые опираются на птеригиофоры (Ү. Franҫois, 1955, 1957). У кеты и других тихоокеанских лососей дифференцировка анального плавника несколько опережает дифференцировку спинного. Постепенно разрастается брюшная лопасть хвостового плавника. В нем формируется до 8—10 лучей. Нижние остистые отростки последних позвонков образуют широкие пластинкигипуралии.
В течение этапа густота капиллярной сети желточного мешка продолжает возрастать за счет увеличения числа анастомозов между сосудами. Краевая вена опускается на вентральную сторону желточного мешка. Капиллярные сосуды развиваются почти на всей верхней поверхности головы. Появляются капилляры в зачатках псевдобранхий. Подключичная артерия — а. subclavia образует кольцевые сосуды грудных плавников. Во всех хвостовых миотомах появляются сегментальные сосуды. От передних туловищных сегментальных артерий ответвляются сосуды, идущие к спинному мозгу, в толщу мышечной ткани и к поверхности
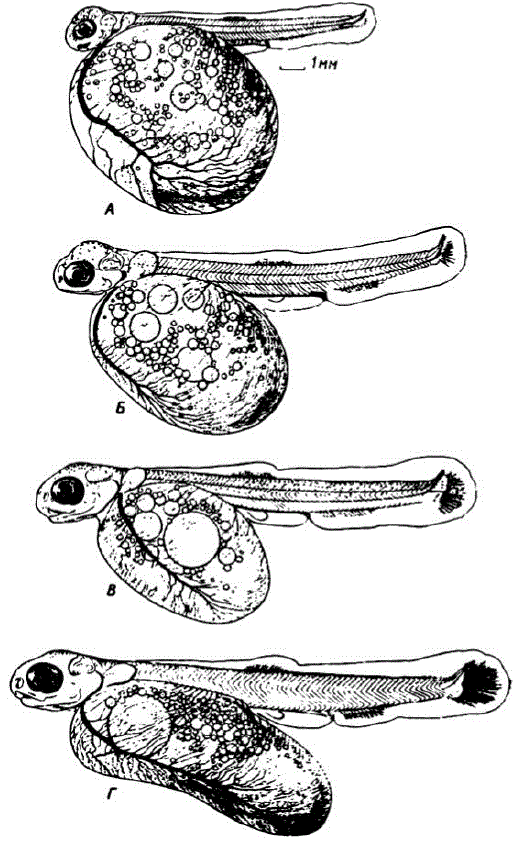
Рис. 11. Позднне эмбрионы осенней кеты A — возраст 30 сут., длина 13,1 мм. Б — возраст 38 сут., длина 17,7 мм. В — возраст 48 суток, длнна 21,5 мм. Г. — зародыш в момент массового вылупления, позраст 51 сут., длина 23,5 мм. Развитие при температуре 92—10.5" (в Соколопском инкубаторе). Прижизненная +присгTкіі
тела. В покровах тела образуется сеть капилляров, несущая дыхательную функцию (Смирнов, 1953б). Ответвления сегментальных сосудов над миотомами образуют truпcus longitudinalis dorsalis, у поверхности тела вдоль средней линии — tгuпcus longitudinalis lateralis, они же питают кровью капилляры, проходящие у поверхности тела по межсегментальным миосептам. В началс этапа в покровах головы, а затем и туловища появляются меланофоры.
Развитие верхних и нижних конусов миотомов идет в направлении от хвоста к голове, в порядке, обратном возникновению миотомов. В такой же последовательности дифференцируются и позвонки. Более раннес усложнение формы миотомов хвоста А. Г. Рындзюнский (1939) связывает с его важностыo для плавания, т. е. с функцией органа на болес поздних этапах развития. Следуст замстить, что движения хвоста важны для жизни эмбриона. Они благоприятствуют циркуляции крови и обеспечивают перемешивание перивителлиновой жидкости,
К концу этапа число хвостовых сегментов сокращается до 22—23 и их общее число уменьшается до 65—63. Непарная плавниковая складка в месте закладки непарных плавников расширястся и в них формируются опорные элементы. В хвостовой лопасти развивается довольно густая сеть капилляров. Персдний край нижней челюсти достигает вертикали, опущенной от переднего края глаз. Выявляются контуры костных элементов верхней и нижнсй челюстей. Рот приобретает подвижность. Жаберные крышки прикрывают 3-ю пару жабер. Печень развивается в мощпый орган, пронизапный густой сетью кровеносных сосудов. Заканчивается дифференцировка верхних и нижних конусов миотомов. Обонятсльные ямки приобретают удлиненную форму. Слуховые пузырьки принимают вид неправильных пирамидок. В покровах головы появляются мелкие оранжево-красные липофоры. По бокам головы и под ней на передней части желточного мсшка обозначаются железки вылупления (рис. 11, B). К концу этапа длина зародышей увеличивается до 16—18 мм.
11 этап. Развитие подвижности челюстей, жаберных крышек, завершение инкубации. Настоящим этапом завершается развитие эмбрионов под оболочкой. Длительность инкубации при разной температуре варьирует в широких пределах. В описываемой серии инкубация икринок длилась 51—55 сут. (521—530 градусодней). Основная масса зародышей при вылуплении имела длину 22—23 мм, а некоторые достигали 24 мм. На Тепловском заводе Н. Н. Дислер отметил вылупление кеты в возрасте 122—128 сут. (408—420 градусо-дней), при длине зародышей 20—22 мм. Меньший размер этих зародышей связан с тем, что кета тепловского стада выметывает более мелкие икринки.
В течение этапа продолжает формироваться ротовой аппарат. Нижняя челюсть вырастает за вертикаль переднего края глаз. Рот принимает полунижнее положение. Прорывается дно ротовой воронки. Жаберные крышки прикрывают все жабры. Намечается образование перемычки, разделяющей отверстие обонятельной ямки папереднее и заднее (рис. 11, Г). Рот, жаберные крышки и жаберный аппарат приобретают подвижпость. Движения челюстей и жаберных крышек в икринке затрудняются тем, что крупная голова эмбриона оказывается вдавленной в желточный мешок.
Ко времени вылупления разрастаются лопасти грудных плавников и их вершины достигают 5—6-го миотомов. В спинном и анальном плавниках четко выявляются лучи. Плавники вычленяются из непарной плавниковой складки, которая заметно рсзорбируется. В месте закладки жирового плавника намечается скопление мелких меланофоров и липофоров. Вершины увеличив1шихся брюшных плавников приближаются к краю преанальной плавниковой складки. В хвостовом плавнике развивается 15—19 лепидотрихий.
Дорсальная поверхность зародышей заметно темнеет. Зерна пигмента в хроматофорах приобрели способность мигрировать, благодаря чему зародыши могут изменять окраску. На туловище меланофоры многочисленны до средней линии тела, а ниже единичны. Появились мсланофоры между лучами хвостового плавника. Образуется ряд пигментных клеток над спинным мозгом (невральный ряд), вдоль крупных кровеносных сосудов (гемальный ряд), пигментируется выстилка брюшной полости. Мелкие красно-оранжевые липофоры покрывают дорсальную поверхность головы и в небольшом количестве появляются на спине зародыша. Позади зачатков спинного и анального плавников липофоры пропикают в основание непарной плавниковой складки.
К концу этапа капиллярная сеть на желточном мешке достигает максимальной густоты. Длина капилляров на 1 мм2 его поверхности достигает 7,5-8 мм. Усложняется сеть капилляров вблизи поверхности тела. Нужно заметить, что при наркотизации зародышей мелкие капилляры покровов тела быстро запустевают, и эти сосуды следуст изучать без применения паркоза.
За несколько дней до вылупления в гопадах выделяются крупные зародышевые клетки диаметром 7,2—15,4 мк. В их ядрах наблюдаются фигуры митоза. Митотическое деление первичных половых клеток продолжается и после вылупления (Robertson, 1953).
Эмбрионы способны производить хвостом энергичные волнообразные движения и при возбуждении начинают вращаться под оболочкой. Если такого зародыша извлечь из икринки, он способен подняться в толщу воды и продвинуться на несколько сантиметров вперед, но локомоции хвоста быстро ослабевают и эмбрион опускается на дно. При благоприятных условиях аэрации эмбрионы в икринке движутся редко, что помогает экономить запасы энергии. Перемешивание перивителлиновой жидкости в это время осуществляется в основном благодаря ритмичному движению грудных плавничков. Выполнению той же задачи служат движения рта и жаберногоаппарата.
К концу инкубации весь мочеточник и мочевой пузырь заполняются накопленными и кристаллизовавшимися продуктами азотистого обмена. Как известно, во время инкубации икры через оболочку выделяется лишь небольшое количество аммиака (Светлов, 1928; Hayes, 1949). Для лососей, откладывающих икру кучно в грунт, где проточность слабая, отмеченная особенность имеет жизненно важное значение, так как благодаря этому не загрязняется вода нерестовых бугров.
Непосредственно перед вылуплением эмбрионы ведут себя беспокойно, энергично ворочаются. Под действием протеолитического фермепта, выделяемого железками вылупления, яйцевые оболочки разрушаются и эмбрионы выходят наружу. Опыты на икре европейского лосося показали, что фермент вылупления выделяется в подоболочечное пространство за короткое время и быстро разрушает оболочку. Подготавливает оболочку к действию фермента вылупления накопившаяся гиалуронидаза. Значение механических факторов в акте вылупления невелико. В опытах, обездвиженные наркозом зародыши вылупляются вполне успешно исключительно за счет растворения яйцевых оболочек ферментом вылупления. Адекватным раздражителем, возбуждающим секреторную деятельность желез вылупления, является снижение парциального давления кислорода в перивителлиновой жидкости. Процесс секреции протекает, возможно, как нейрогуморальный рефлекс (Бузников, 1955, 1961, 1964; Бузников, Игнатьева, 1958). «Вылупление может произойти или раньше, или позже по отношению к развитию самого зародыша в зависимости от влияния условнй среды» (Васнецов, 1953а, стр. 210).
Поскольку железки вылупления у лососей сгруппированы на голове и передне-верхнем участке желточного мешка, выделяемый ими секрет мог бы действовать локально. Это предотвращается движениями зародышей, с помощью которых фермент распределяется в подоболочечном пространстве и воздействует на всю оболочку. Если же складываются неблагоприятные условия, зародыши угнетаются и утрачивают подвижность, секрет желез вылупления действует локально, растворяет небольшой участок оболочки и наружу высовываются только головы, выход зародышей затрудняется и они могут погибнуть. В условиях высокого насыщения воды кислородом вылупление может задерживаться; к этому же приводит и излишне сильный ток воды. Рыбоводы заинтересованы в быстром прохождении вылупления. «Дружный выход зародышей из оболочек можно получить только в условиях слабого тока воды, т. е. в условиях, которые приближаются к естественным» (Дислер, 1957, стр. 27).
На рыбоводных заводах разных страп наблюдались заболевания икры, приводящие к преждевременному ослаблению яйцевых оболочек, их разрушению и выходу ослабленных эмбрионов на ненормально ранних стадиях развития. Подобное заболевание могут вызывать грамотрицательные бактерии, некоторые простейшие животные, одноклеточные грибки из класса Агchymyceteae. Иногда заболевание принимает чрезвычайно большие масштабы, и гибнут десятки миллионов икринок (Напda, 1933, 1934; Ляйман, 1957; Сакун, Чистова, 1960). Заболевание удается предупреждать и успешно ликвидировать, применяя для профилактики и лечения формалин, малахитовый зеленый и другие препараты, о чем свидетельствует практика Соколовского и других сахалинских заводов (Смирнов, 1963б, 1964б).
Еще В. К. Солдатов (1912) отмечал, что зародыши выходят в разные сроки из икринок, взятых даже от одной самки, и связывал это с индивидуальными особенностями икринок. В условиях Соколовского завода массовое вылупление длится 2—3 дня, но может растягиваться на 7—10 дней и более, если температура воды низкая. На том же заводе, в зависимости от температуры, сроки инкубации икры колеблются от 49 до 80 и более суток, но чаще вылупление приходилось на 53—60-е сут. На других заводах, приспособленных для разведения кеты, сроки инкубации варьируют от 45 до 196 суток (223—610 градусо-дней). В одних случаях вылупление начинается в конце октября (Соколовский завод), в других — весной (Краснопольский завод) и продолжается, соответственно, до ноября или мая. Сравнение показывает большие различия темпа развития и сроков инкубации кеты на Соколовском и Тепловском заводах; подобные различия наблюдаются и в природе. Замечено, что с понижением температуры воды время инкубации икры увеличивается, тогда как число градусо-дней уменьшается. Поэтому определение темпа развития в градусо-днях имеет орнентировочное значение.
12 этап. Пассивное состояние свободных зародышей. Этап начинается после выхода зародышей из оболочек и длится при температуре 3,3° 5—7 дней, при температуре 9" — 3—4 дня. Вылупившиеся зародыши слабо реагируют на внешние раздражители и мало подвижны. Их длина варьирует в широких пределах. Размер зависит от величины икры и стадии развития. Так, из икринок среднего диаметра 8,7 мм зародыши выходили длиной 21,0—24,5 мм, при модальной длине 22,5—23,0 мм, тогда как из икринок диаметром 7,9 мм вылуплялись зародыши длиной 19— 23 мм, при модальной длине 20,5—20,7 мм. Мелкие экземпляры, выходящие из икринок сходного размера, имели в хвосте на 1—3 луча меньше, нежели крупные зародыши. На Поронайском заводе кета из смешанной икры вылуплялась при длине 21 — 25 мм (М=23,0+0,08) и весе от 149 до 240 мг, причем М = 1996-2,1 мг (Житенева, Галкина, 1965). IIa. Тепловском за. воде зародыши выходят из икринок, имея длину от 17,3 до 22,0 мм, в среднем 19,0 мм, и вес 174 мг (Леванидов, 1954б; Дислер, 1957). Различия объясняются небольшим размером икринок кеты TeIIIOBCKOTO CTaLa.
При выходе из икринки кета имеет огромный желточный мешок яйцевидной формы. В желтке содержится много мелких жировых капель и обычно одна крупная, диаметром около 3 мм. При изменении положения зародыша жировые капли в желтке по-прежнему могут перемещаться. На рис. 11, Г изображен зародыш, у которого за время зарисовки жировые капли сместились на левую сторону.
Число туловищных сегментов осталось прежним, в хвосте к моменту вылупления оно сократилось до 22—21. Хвостовые составляла 36,5% всей длины зародыша. Форма хвостовой лопасти асимметрична.
Непарная плавниковая складка продолжаст резорбироваться. Из нее обособился спинной плавник, наметились контуры жирового плавничка. В спинном и анальном плавниках заложилось большинство лепидотрихий. Ко времени массового вылупления в хвостовом плавнике обычно закладывается 17—19 лепидотрихий. Если же при вылуплении их количество менее 15, это указывает на раннее вылупление. У зародышей образовался костный плечевой пояс. Грудные плавники крупные, округлой формы. В брюшных плавниках опорные элементы еще отсутствуют. Преанальная плавниковая складка ко времени вылупления достигает максимального развития.
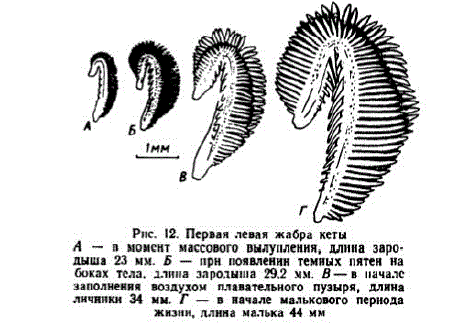
Рис. 12. Первая левая жабракеты A — п момент массового вылупления, длина зародыша 23 мм. Б — прн появленин темных пятен на боках тела, длина зародыша 29,2 мм. В — в началс заполнення воздухом плавательного пузыря, длина личинки 34 мм. Г. — в начале малькового пернода жизни, длина малька 44 мм
Роту вылупившихся зародышей занимает полунижнее положение. В слуховых капсулах четко различаются полукружные каналы и крупные отолиты. Статоакустические органы максимально сблизились с глазами. Жаберные крышки закрывают жабры.
На первой жабре средние лепестки имеют длину 0,25 мм и становятся ветвистыми; в лепестках появились капиллярные сосуды. Закладываются жаберные тычинки, имеющие вид небольших бугорков (рис. 12, A).
В начале этапа желудочно-кишечный тракт представляет собой почти прямую трубку. Зачаток желудка, имеющий длину около 1 мм, выделяется утолщенными стенками и некоторым
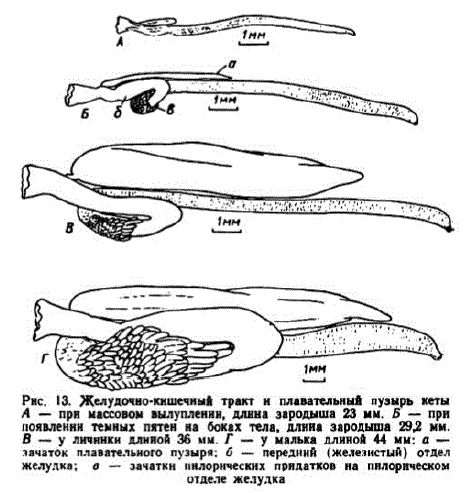
Pнс. 13. Желудочно-кншечный тракт н плавательный пузырь кеты
A — при массовом вылуплении, длина зародыша 23 мм. Б - при
появленнн темных пятен на боках тела, длина зародыша 292 мм.
В — уличннки длиной 36 мм. Г. — у малька длиной 44 мм: а —
лачаток плавательного пузыря; б — передний (железистый) отдел
желудка, о — зачаткн пнлорических прндатков на пнлорическом отделе желудка
сужением на границе с кишкой. На его будущем пилорическом отделе заметны почкообразные выпячивания — зачатки первых пилорических придатков. Длина кишки 7,5—8,0 мм — около 39% общей длины зародыша. Слизистая кишки приобрела складчатость. Зачаток плавательного пузыря вырос до 1,1—1,2 мм
(рис. 13, A).
Мочевой пузырь после вылупления начинает периодически опорожняться и заполняться вновь. Вместе с этим уменьшается количество кристаллов, обильно заполнявших мочеточник. Зародыши быстро освобождаются от накопленных продуктов обмена
63
и если в питомниках проточность слабая, а плотность посадки чрезмерно высокая, окружающая зародышей среда может ухуд
LIHTLORI.
Ко времени вылупления формируются темные пятна над глазами и слуховыми капсулами, меланофоры равномерно покрывают всю дорсальную половину тела, располагаются между лучами хвостового плавника. Усиливается и липофоровая пигментация головы. Липофоры появляются на всей дорсальной стороне тела и в хвостовом участке непарной плавниковой.
Органы боковой линии — невромасты, — имеют вид бугорков. они образуют ряды: падглазничный, подглазничный, височный и преоперкуло-мандибулярный; появляются невромасты и около обонятельных отверстий. Вдоль средней линии тела чувствующие бугорки дифференцировались до середины хвостового отдела или нcмного дальше. Невромасты надвисочной и югальной групп еще не дифференцированы. Чувствующие бугорки у живых зародышей прокрашиваются водным раствором метиленовой сини, нервы же, иннервирующие их, еще не прокрашиваются, что указывает на недостаточную функциональную зрелость (Дислер, 1957, 1960). Выйдя из оболочек, зародыши остаются лежать между камнями нерестовых бугров. В питомниках с ровным дном они ложатся на бок. Грудные плавнички находятся печти в постоянном движенин, что способствует смене воды у поверхности желточного мешка и тела и имеет дыхательное значение. Ритмичная работарта и жаберных крышек обеспечивает хорошую вентиляцию жабер. Потребление кислорода после выхода эмбрионов из оболочки резко возрастает.
В питомниках зародыши временами начинают активно двигаться, переворачиваются спиной кверху и устремляются вперед. Движутся они без отрыва ото дна, наклонив голову книзу. Будучи помещенными в толщу воды, быстро опускаются ко дну. Их маневренности мешает большой желточный мешок. Опыты показали, что такие зародыши на струи воды реагируют еще не четко, к прикосновению каким-либо предметом относятся безразлично, на стук не реагируют. На свет зародыши реагируют слабо отрицательно и при ярком освещении постепенно передвигаются в затененные участки (Дислер, 1957, 1960). В гнездах вылупившиеся зародыши некоторое время остаются там, где лежала икра, а затем начинают передвигаться по щелям между камнями. В зависимости от режима водоснабжения нерестилищ опи могут двигаться в разные стороны, со временем приближаются к поверхности групта. В питомниках вылупившиеся зародыши проваливаются через ячею рыбоводных рамок и скапливаются под стопками. Вследствие большой скученности, они закрывают друг другу поверхность желточного мешка, важную для дыхания, и могут задохнуться даже при хорошем насыщении воды кислородом. Такие скопления надо своевременно рассеивать.
укрываться от света, активно двигаются навстречу течению. Появляется ярко выраженная положительная реакция на прикосновение посторонних предметов (положительное тактильное чувство). На Тепловском заводе в начале этапа зародыши имели длину 24—26 мм. При температуре 3,4° этап длился 45—48 сут. На Соколовском заводе при 8,5° этап длился 25—27 сут.
В течение этапа формируются непарные и брюшныс плавники. Очень быстро растет плавательный пузырь, удлиняется и дифференцируется пищеварительный тракт. К концу этапа личинки достигают длины 30—32 мм и начинают потреблять внешнюю пищу.
Ускоренно растет хвостовой отдел и через 20 сут. после вылупления длина постанального отдела превышает 38,5% общей длины. Продолжается резорбция дорсальной и вентральной плавниковых складок. Преанальная складка сохраняет прежний размер. Увеличивается высота спинного, анального и жирового плавников. В спинном и анальном плавниках закладываются всcлучи. Хвост снова приобретает симметричную форму. Ускоренный рост верхних и нижних лучей приводит к образованию вырезки лопасти хвостового плавника. В его крупных лучах формируется до 3—4 члеников. Закладываются лучи в грудных и брюшных плавΗΗΗΗΜ :
На чcлюстях появляются зачатки зубов, которые прорезаются к концу этапа. По краям жаберных крышск вырастают кожистые складки и в них формируются лучи. Кожистые складки достигают плечевого пояса и хорошо прикрывают жабры. Нижние челюсти и жаберные крышки совершают ритмичные дыхательные движения. Смене воды в жаберной полости способствует развившийся во рту парус — veluп. Смыкаются боковыс выросты стенок обонятельных ямок, обособив переднес и заднее обонятельные отверстня. Расстоянне между глазами и слуховыми капсулами начало медленно увеличиваться. Возрастает абсолютный и относительный размер головы.
После вылупления зародышей резорбция желтка ускоряется. При изучении кеты и других лососей показана важная роль в этом процессе перибласта, входящего в тесный контакт с желточными венами в мсстах отсутствия у них эндотелня (Согdiег, 1941; Yamada, 1959). Со временем все большая часть желточного мешка закрывается вентральными выростами миотомов, мелкис кровеносные сосуды на его поверхности запустевают и все большую роль в обеспечении организма кислородом играют жабры. Дыхательную функцию несут также капилляры, расположенные в лопасти хвостового плавника и у поверхпости тела.
За счет формирующихся изгибов удлиняется желудочнокишечный тракт. Кишечник заполняется содержимым зеленоватожелтого цвета, заметны перистальтические движения его стенок. Увеличивается число пнлорических придатков и их размер. Быстро растет плавательный пузырь (рис. 13, Б). Васкуляризуется средняя область гонад и продолжается размножение зародыше. вых клеток (Robertson, 1953).
Mсланофоры по бокам тела распространяются до нижних конусов миотомов, появляются в основании и вдоль переднего края спинного плавника, их становится заметно больше в хвосто. вом плавнике. Усиливастся пигментация оболочек головного и спинного мозга, выстилки брюшной полости, кровеносных сосудов. Мелкие красно-оранжевые липофоры распространяются ниже средней линии тела.
Заканчивается дифференцировка чувствующих бугорков оргапов боковой линии па голове и по всему телу. Водным раствором метиленовой синн прокрашиваются не только невромасты, но и нервы их иннервирующие, что указывает на функциональную зрелость этой системы органов чувств. Зародыши четко реагируют на ток воды и движутся навстречу ему, способны противостоять слабому течению. Они воспринимают токи воды, вызываемые движением других особей, и вслед за возбуждением нескольких экземпляров в движение приходит вся масса зародышей. Ярко выражена положительная реакция на прикосновение. Эта реакция возникла в связи со своеобразием каменисто-песчаных гнезд и препятствует преждевременному выходу свободных зародышей из грунта. Оказавшись по каким-либо причинам вне гнезда, зародыши возбуждаются и активно плавают до тех пор, пока опять не попадут в расщелины между камнями. Преждевременному выходу личинок из гнезд препятствует также их светобоязнь.
Как известно, солнечный свет на икру и вылупившихся зародышей лососей действует отрицательно. По наблюдениям Н. Н. Дислера (1957), при средней температуре 4,1° пагубнос действие света отмечается до одномесячного возраста после вылупления. Солнечный свет вызывает обесцвечивание эритроцитов и уменьшение их количества. Под его действием интенсивнее развивается черный пигмент, тогда как оранжсвый разрушается и сохраняется только на теменной поверхности головы. На желточном мешке появляются белесые островки отмирающей ткани. К разрушенным участкам присасываются планарии, которые поглощают желток и ускоряют гибель зародышей. Изменение окраски рыб под воздействием света отмечалось и раньше. Было показано участие каротнноидов в светочувствительных реакциях pl6 (Sumner, Fox, 1933, 1935).
С возникновеннем положительной реакции на течение, прикосновение к посторонним предметам, светобоязни зародыши в питомниках с ровным дном начинают двигаться навстречу притока воды и ищут укрытий. Они скапливаются у мест поступления воды, в углублениях дна, забиваются в щели между досками. Подобные скопления, как говорилось, опасны для жизни зародышей и на их разгонку рыбоводы вынуждены тратить много усилий. Во время перемещений в питомниках зародыши о шероховатое дно повреждают нежный покров желточного мешка, что может привести к гибели. В питомниках иногда наблюдается дугообразное искривление туловища, изменение формы желточного мешка, смещение жировой капли пазад, где она может давить на кишечник. Такие изменения связываются с выдерживанием свободных зародышей кеты на ровном дне, то есть в несвойственных для них условиях. Указывается, правда, на нивелировку подобных отклонений в ходе дальнейшего развития. В связи с освещенностью питомников ускоряется развитие предметного видения, кета несколько раньше переходит на плав и может преждевременно покидать питомник (Дислер, 1954, 1957, 1960). Естественным потребностям свободных зародышей отвечает покрытие питомников несколькими слоями крупной обкатанной гальки, которая должна омываться водой снизу.
Этап завершает зародышевый период развития кеты. На Тепловском заводе он заканчивался в возрасте 177—180 сут. (601— 612 градусо-дней), на Соколовском заводе в возрасте 80—83 сут. (число градусо-дней оказывалось значительно большим —
760-788).
Смотрите также
Бассейновый метод выращивания лососевых рыб