Развитие кеты
Представляется целесообразным сделать некоторые замечания, касающиеся периодизации оптогенеза и терминологии, прпменяемой при описании развития, поскольку в литературе по этим вопросам встречаются расхождения.
В морфологии под стадией развития принято понимать любое произвольно взятое состояние организма (Васнецов, 1953а; Крыжановский п др., 1953). В качестве примеров можно назвать стадии 2-, 32-бластомеров, стадии 5, 20 сегментов, эмбрион 6 мм длины, стадию начала пигментации глаз и т. д. Совокупность последовательных стадий определепного интервала онтогенеза, в течение которого развивающийся организм и его связи со средой существенно не меняются, В. В. Васнецов назвал этапом развития. «Каждый этап отличается от других специфическим для него строением, физиологией и биологией, причем и строение, и физиология приспособлены к данной биологии и данным специфическим условиям» (Васнецов, 1953а, стр. 210). Мы будем придерживаться указанного содержания этих понятий. Разными авторами этапы обозначаются буквами, цифрами, называются по основным морфофункциональным признакам и особенностям экологии.
С. Г. Крыжановский обратил внимание на то, что в течение каждого этапа происходят и количественные и качественные, и
постепенныс н скачкообразные изменения, в прсделах опрсделсIIной меры нс изменяющие, однако, основного качества, характеризующего этап. Только после достижения меры количество переходит в новое основнос качество, организм переходит на новый этап развития. Все основные предпосылки для перехода в новое качество, на новый этап развития создаются па предшествующем этапе развития (Крыжановский и др., 1953, стр. 72).
Группы этапов, характеризующиеся общими приспособлениями, в свою очередь, объединяются в периоды развития.
В эмбриональный период онтогенеза зародыши питаются матерпнскими запасами пищи, заключенными в желтке. Такое эпдогcпное питание свойственно не только зародышам, находящимся под оболочкой икринок, или собственно эмбриопам, но сохрапяется определенное время и после их вылупления у свободных зародышей. По этой причине данный интервал онтогенеза можно также называть зародышевым псриодом, или псриодом эндогепΗΟΓΟ ΠΗΤΕHΗΗ.
Важнейшее биологическое отличие личинок состоит в переходе к активному добыванию пищи из внешней среды и наличии провизорных личиночных органов. Задаче активного добывания пищи подчинены приобретаемые личинками морфологические, физиологические и экологические особенности, сохраняющиеся в течение личиночного периода развития. Лососи подготавливаются к началу активного питания, имея сще большой остаток желтка, и для их личинок характерен продолжительный срок смешанного эндогенно-экзогcнного питания. При отсутствии доступной пищи время смешанного питания сокращается.
К мальковому периоду жизни провизорные личиночные органы исчезают и появляются некоторые характерные для взрослых рыб органы (например, непарные и парные плавники со скелетом и мускулатурой, чешуя). Соответственно меняется среда обнтаIIII H II OBCEPHHC,
Существенная особенность жизнедеятельности взрослых рыб состоит в интепсивном нагуле, обсспечивающем своевременное созревание половых желез и успешное размножcние. Нагул лососи могут осуществлять как в пресной воде (жилые и остаточные формы), так и в морской (анадромные, или проходные, формы).
Выходу лососей на места икрометания предшествуют глубокие морфофункциональные превращения. Во время нереста меняют ся среда и поведение. Комплекс разносторонних изменений свидетельствует о биологической специфичности этого интервала онтогенеза, выделенного нами в нерестовый этап, который вместе с преднерестовой миграцией логично объединить в период размножения (Смирнов, 1959б, 1964в, 1965; Еремеева, Смирнов, 1965; , 1967). После размножения тихоокеанские лососи погибают.
Переходим к характеристике половых продуктов и развития кеты.
оловые продукты. Сперматозоиды кеты, так же как у родственных ей видов и других костнстых рыб, жгутикового типа (Riddle, 1917; Lowman, 1953; Гинзбург, 1968). Созревшие сперматозоиды лососи выделяют отдельными порциями в течение продолжительного времени. Чтобы составнть представление о сроках и количестве выделяемой спермы, зрелых самцов выдерживали в садках (Смирнов, 1963а, 1964б). По мере созревания очередной порции сперма отцеживалась прижизненно. Определялся объем отдельных порций спермы (объем эякулята), концентрация сперматозоидов, способность спермы оплодотворять икру.
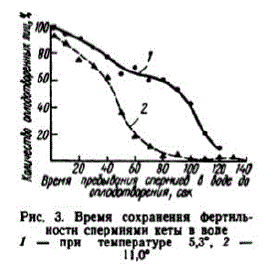
Отдельные самцы выделяли сперму почти в течение месяца. С перерывами в 1—2 или 3 суток от одного самца удавалось получить до 10—11 порций спермы. Объем эякулята варьировал от 3,6 до 17,9 мл и составил в среднем 9,2 мл. От одного подопытного самца было получено свыше 130 мл спермы. В наиболее густой сперме в 1 мм3 содержалось 32,4 млн. сперматозоидов, тогРис. 3. Время сохранення фертнль- да как в жидкой — 17,36 млн.;
ностнспермнями кеты в вопе средняя величина - 24,12 млн. / — npH Tempt * Triocenokaa. To
в эякуляте среднего объема содержится более 220 млрд. сперматозоидов. Сперма разных порций использовалась для искусственного оплодотворения икры. Проверка показала ее доброкачественность,
При температуре 10—11° энергичное поступательное движение сперматозоидов осенней кеты и горбуши в воде, взятой с мест нереста, наблюдалось в течение 30—55 сек.; слабые колебательные движения отмечались через 90 и даже 170 сек. Это близко к тем срокам, которые указываются для озерной форели (Гинзбург, 1968).
Попав в воду, спермии быстро утрачивают фертильность (рис. 3). В наших опытах при температуре 11" уже через 10 сек. разведенная сперма оплодотворяла икринок на 6% меньше, чем при «сухом» способе осеменения, через 30 сек. — на 22% меньше, а после 90 сек. разбавленной водой спермой оплодотворялись единичные икринки. При более низкой температуре сперматозоиды сохраняли фертильность дольше.
Кета выметывает очень крупные икринки. У проанализированных 64 самок осенней кеты из р. Б. Такой средний вес зрелых яиц колебался от 183 до 300 мг (М=239,25+0,19; o=29,8). Диаметр набухших икринок осенней сахалинской кеты колеблется от 6.7 до 9,0 мм, модальная величина — 7—8 мм. Яйца телолециталь
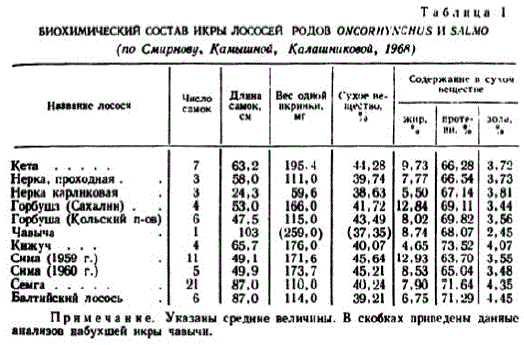
Примечанне. Указаны средние величины. В скобках приведены данные анализов набухшейнкры чавычн.
У лососевых рыб яйцевые оболочки состоят из лучистой зоны (zопа гadiata) и тонкого внешнего гомогенного слоя. Толщина оболочек набухших яиц осенней кеты около 55 мк (55,1+0,38), Построена оболочка из ихтулокератина (Bell et al., 1969). У кеты дифференцируются два слоя лучистой зоны (К. Yaпапоto, 1951; Капоh, Yaпапоto, 1957; собствен. данные). Внутреняя часть лучистой зоны, толщиной около 5 мк, отличается болес рыхлой струк೪ಕ್ಕಿ Гомогенпый поверхностный слой нмеет толщину 1—1,5 мк. S. gairdпегі (Rich.) наружный слой хориона содержит нейтральные карбогидраты, белок и мукополисахариды несульфатного типа (Hagenmaier, 1973). Он предохраняст икру от действия на пее ферментов вылупления с внешней стороны (Bell, Hoskins, Begshaw, 1969).
У осенней кеты икринки ярко-оранжевые. Сравнение цвета нкры разных самок с цветовыми полями Атласа цвстов (Рабкин, 1956) позволяет охарактеризовать вариации цветового тона длиной волны 589—592 ммк. Окраска обусловлена каротиноидным пигиентом. Оплодотворенные икрипки камчатской кеты содержат 1,60—l,76% acTaKcâHTHHa (5HpxKoM6eK, 1970).
Время, в течение которого яйца кеты в воде сохраняют спо
собность к оплодотворению (фертильность), указывается от 2 до 30 мин. (Kusa, 1950, K. Yaпапоto, 1951). В связи с этим важно подчеркнуть, что основная масса икринок после активацин водой быстро утрачивает фертильность. В наших опытах при температуре 10—11° через 1 мин. после выдерживания икринок в воде оплодотворялось на 40% меньше, нежели при «сухом» способе оссменения (контроль), через 3 мин. — на 80—90% меньше, а через 12 мин. оплодотворялись лишь единичные икринки (рис. 4).
 Рис. 4, Время сохранення фертиль- Рис. 5. Температурный режим ности яйцами кеты п воле ннкубаторов в годы сбора матеI — при температуре
Рис. 4, Время сохранення фертиль- Рис. 5. Температурный режим ности яйцами кеты п воле ннкубаторов в годы сбора матеI — при температуре
Условия развития изучавшегося материала. Описание основывается на наблюдениях за икрой, развивавшейся в инкубаторе Соколовского завода. В инкубатор подавалась чистая ключевая вода, содержащая кислорода 3,4—8,0 мг/л, свободной углекислоты 20—30, иногда около 40 мг/л, значение pH варьировало в пределах 6,2—6,6. Суточные колебания температуры не превышали 0,1—0,2° (рис. 5). Вода медленно прогревалась летом и в начале осени, в середине октября температура воды достигала 10—10,5°, и только после этого начинала медленно охлаждаться; в декабре температура была 7—8°, наиболее низкая — 2,5—5° — в марте— апреле; в конце апреля вода вновь начинала прогреваться.
Темп развития кеты на Соколовском заводе мы сравниваем с темпом развития амурской осенней кеты на Тепловском заводе (Дислер, 1957). На Тепловском заводе среднемесячная температура воды в 1950 и 1951 гг. составляла, соответственно: в октябре 3,9 и 4,4°, в январе она снижалась до 2,7 и 3,3", в апреле прогревалась до 5°, а в мае — до 6,8°. Насыщение кислородом воды в аппаратах составляло 40—60% от нормального, а количество углекислоты колебалось от 4,4 до 17,0 мг/л.
Смотрите также
Бассейновый метод выращивания лососевых рыб
Биология, размножение и развитие тихоокеанских лососей
Атлантический лосось на севере России