Факторы, определяющие численость покатной молоди кеты(ONCORHYNCHUS KETA) Р. Иска
Исследованию причин варьирования урожайности поколений животных и растений, особенно тех видов, которые играют заметную роль в жизни человека, всегда уделялось большое внимание. Живые организмы наиболее уязвимы на самых ранних стадиях жизненного цикла, когда погибает наибольшая часть поколения, поэтому незначительные вариации факторов, влияющих на выживаемость потомков в это время, могут повлечь как депрессию, так и вспышку численности поколения. Это определяет важность исследования факторов, влияющих на численность поколений лососей в начальном, пресноводном, периоде жизни.
Основная цель работы — анализ причин изменчивости численности покат-ной молоди кеты р. Иска. Учитывая множественность причин вариации численности потомков, вполне очевидно, что приблизиться к пониманию данного явления невозможно без применения методов имитационного моделирования. В процессе построения модели неизбежно появляются ее варианты, выбор окончательного варианта не всегда можно строго обосновать, используя лишь формальные критерии. Дополнительная цель исследования — определить, как изменяется оценка роли факторов, влияющих на зависимую переменную, в связи с выбором варианта модели.
Материалы и методы
Работа основана на анализе результатов сплошного учета численности родителей кеты, прошедших через рыбоучетное заграждение в р. Иска (Сахалинский залив, Охотское море), и выборочного учета численности покатной молоди с использованием мальковых ловушек Таранца в период 1970-1998 гг. Материалы ежегодно предоставлялись ФГУ “Амуррыбвод” для разработки прогноза запаса лососей. Кроме того, в работе использованы доступные сведения (© thermograph.ru) по среднегодовым и среднемесячным значениям минимальных, максимальных и средних температур воздуха, годовому и ежемесячному количеству осадков по данным ближайшего от р. Иска метеорологического поста г. Николаевск-на-Амуре.
Методика множественного регрессионного анализа описана в справочной литературе (Поллард, 1982; Дрейпер, Смит, 2007; Медик, Токмачев, 2007; и др.). Усложнение модели за счет ее дополнения новыми факторами прекращали, если оно приводило к значительному увеличению ошибок коэффициентов, когда 95 %-ные доверительные границы средних значений коэффициентов, построенные по их асимптотическим стандартным ошибкам, включали значение 0 (за исключением свободного члена в линейных уравнениях). Для уточнения ошибок коэффициентов модели и анализа ее устойчивости использовали 100 бутстреп-выборок с использованием остатков.
В уравнениях прописными буквами обозначена численность (в тысячах экземпляров): Р — родителей кеты; Р£ — производителей горбуши; S, Sf — потомков кеты (соответственно расчетная и фактическая), а также переменные №п — количество осадков, мм; Тп — температура воздуха, оС). Цифровые индексы при Р и Т обозначают календарный номер месяца. Строчными буквами (а^) обозначены коэффициенты. В таблицах и тексте использованы следующие обозначения: Р Р^ РТ — уровни значимости соответствующих критериев Е, и Т (Фишера, Шапиро-Вилкса и Стьюдента); d — критерий Дарбина-Уотсона; Б.е., а^.е. — стандартная и асимптотическая стандартная ошибки; Di — разность фактических ^) и расчетных ^) значений численности потомков (остатки), цифровой индекс при D — номер уравнения; Dr, Dc — соответствующие индексам остаточная и общая скорректированная сумма квадратов отклонений; к., кс — соответствующие индексам числа степеней свободы; Р2с — скорректированный коэффициент детерминации; М — среднее арифметическое значение; As, Ех — коэффициенты соответственно асимметрии и эксцесса.
Связи численности потомков кеты р. Иска на стадии миграции в море с численностью родителей и любым из метеорологических факторов слабые. Варьируя способы описания данных связей, не удалось выявить ни одного фактора, влиянием которого можно было бы объяснить более 30 % дисперсии зависимой переменной. Формальный подход к построению модели, в котором очередность включения в нее новых факторов определяется по значениям частных коэффициентов корреляции или Е-критерия Фишера, не дал удовлетворительных результатов — суммарным влиянием всех факторов в различных вариантах модели удавалось описать от 41 до 69 % вариации численности потомков. Лишь два равнозначных варианта линейной модели, построение которых начинали с численности родителей (1а):
оказались приемлемы. Значения параметров обоих вариантов модели и критериальная статистика одинаковы, но коэффициент детерминации в стартовом уравнении 16 больше, поэтому рассмотрим второй вариант модели.
Остатки уравнения 16 (далее — уравнение 1) наиболее хорошо объяснимы изменчивостью численности родителей, а включение данного фактора в модель приводит к значимому (Е = 26,16; РЕ < 0,01) уменьшению дисперсии численности покатной молоди (табл. 1, 2). Суммарным влиянием количества осадков в августе и численности родителей (табл. 1, 3, уравнение 2) объяснимо 63,6 % изменчивости зависимой переменной. Гипотеза о равенстве коэффициентов данного уравнения 0 отвергается с вероятностью более 99 % (Е = 22,67). Хотя вероятность нормального распределения остатков уравнения 2 низка (РШ = 0,031), они не обладают значимой асимметрией или эксцессом (табл. 3), т.е. отклонение распределения от гауссовского можно считать умеренным (Дуброва, 2003). По такой же схеме модель дополняли остальными факторами, последовательность ее усложнения приведена в табл. 1-3.
Зависимость численности потомков от численности родителей слабая (уравнение 1а), она описывает лишь общие тенденции изменчивости численности потомков (рис. 1). В начале и в конце периода исследования численность потомков, рассчитанная на основе данной переменной, как правило, больше фактической, в середине периода — меньше фактической, однако данные отклонения относительно хорошо объяснимы влиянием остальных факторов (табл. 3, рис. 1).
Асимптотические значения стандартных ошибок коэффициентов модели (уравнение 6, табл. 3) варьируют в пределах 9,1-46,3 % от средних значений коэффициентов, в среднем относительная ошибка коэффициентов составляет 20,2 %. Пределы варьирования относительных ошибок коэффициентов, найденных при анализе бутстреп-выборок, значительно уже — 0,8-3,8 % (в среднем 1,8 %). При этом средние значения коэффициентов отличаются от модельных значений лишь на 0,22-2,43 % (в среднем на 1,15 %). Среднее значение критерия Фишера в бутстреп-выборках составило 53,926 ± 2,103, скорректированного коэффициента детерминации — 0,929 ± 0,002. Поскольку модель адекватно описывает данные при варьировании входных параметров в бутстреп-анализе, можно сделать вывод о ее относительной устойчивости.
Значение критерия Дарбина-Уотсона (1,755) превышает верхнее критическое значение = 1,715), т.е. с вероятностью более 99 % можно заключить, что
Таблица 1
Последовательность построения модели, описывающей зависимость численности покатной молоди кеты р. Иска от численности родителей и метеорологических факторов

Таблица 2
Значения коэффициентов и результаты дисперсионного анализа уравнений, описывающих зависимость остатков (Dj—D5, табл. 1) от численности родителей и метеорологических факторов

Примечание. Во всех случаях kc = 28, kr = 27.
сериальная корреляция остатков первого порядка отсутствует. Судя по значению относительной среднеквадратической ошибки модели (34,0 %), ее точность удовлетворительна (Медик, Токмачев, 2007).
Расчетная численность потомков кеты р. Иска при среднемноголетних значениях всех факториальных переменных (стандартные условия), как и среднемноголетняя фактическая численность, равна 3726 тыс. экз. (рис. 2, пунктирная линия). Согласно расчетам, при стандартных условиях, но увеличении на 1 % количества августовских осадков численность покатной молоди увеличивается на 108 тыс. экз. Величина приращения численности потомков при увеличении
Таблица 3
Значения коэффициентов и результаты анализа уравнений 2—6 (табл. 1), описывающих зависимость численности покатной молоди кеты р. Иска от численности родителей и метеорологических факторов
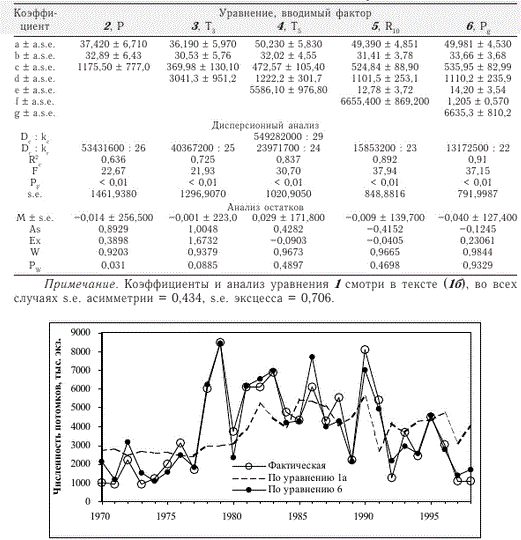
Рис. 1. Динамика эмпирических и расчетных значений численности потомков кеты р. Иска (уравнения см. в тексте и табл. 1, коэффициенты — в табл. 3)
значения единственной факториальной переменной на 1 % и среднемноголетних значениях прочих факторов на рис. 2 указана в скобках.
Модель наименее чувствительна к изменению численности производителей горбуши, нерестившейся за 1 год до нереста кеты. При стандартных условиях прочих факториальных переменных увеличение численности горбуши на 1 % приводит к уменьшению численности покатников кеты на 9,2 тыс. экз. Отклик модели на изменчивость любой из остальных переменных на 1 % при среднемно-
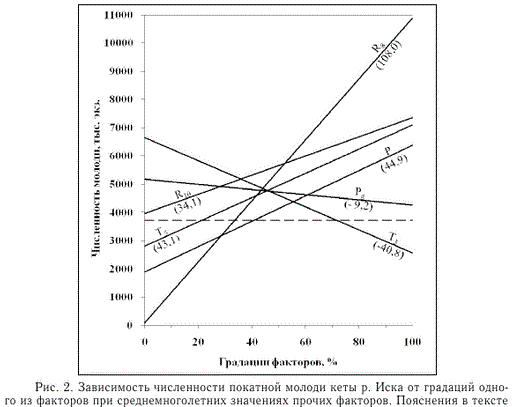
Рис. 2. Зависимость численности покатной молоди кеты р. Иска от градаций одного из факторов при среднемноголетних значениях прочих факторов. Пояснения в тексте голетних значениях прочих факторов изменяется в пределах 34,1-44,9 тыс. экз. (рис. 2).
Таким образом, полученные результаты свидетельствуют об относительно хорошем качестве модели, однако существуют достаточно веские причины полагать, что связь численности родителей лососей с численностью покатной молоди имеет нелинейный характер. Косвенно на это указывает низкая вероятность нормального распределения остатков уравнения 2 (табл. 3). Предположим, что искомая зависимость S(P) (P > 0) должна описываться уравнением S = P • (1 - exp(—(b/P)c)) • exp(a) (7), которое в таком же или несколько измененном виде успешно применялось для описания зависимости численности молоди горбуши и нерки от численности родителей (Островский, Семенчен-ко, 2002; Островский, 2005, 2011; Островский, Пономарев, 2009).
Уравнение 7 оказалось неприменимым ко всем данным, однако связь, описываемая им, достаточно отчетливо проявлялась в начальные годы исследуемого периода. Для анализа причин непостоянства зависимости S(P) использовали метод “скользящего окна”. Начиная с первого года исследования был выделен временной отрезок, достаточный по числу лет для расчета асимптотических ошибок коэффициентов. Минимальная длина временного ряда (“окно”), при которой ошибки коэффициентов были приемлемыми, оказалась равной 13 годам. Фиксированное по длительности “окно”, включающее 13 пар наблюдений “родители— потомки”, смещали по временной шкале с шагом 1 год и для каждого набора данных, попадающих в “окно”, определяли параметры уравнения 7 и критериальную статистику. Результаты данного анализа свидетельствуют, что материалы любого “окна” первых 19 лет действительно могут быть описаны уравнением 7, смещение окна еще на 1 шаг (1977-1989 гг.) приводит к резкому увеличению вероятности равенства коэффициентов уравнения 0 (рис. 3), дальнейшее его смещение усиливает данный эффект.
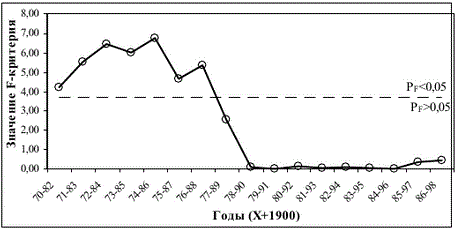
Рис. 3. Результаты проверки гипотезы о равенстве коэффициентов уравнения 7 нулю по F-критерию Фишера методом “скользящего окна”
Все данные первых 19 лет (1970-1988 гг., первый период), так же как данные каждого “окна” (рис. 3), хорошо описываются уравнением 7 (рис. 4, табл. 4-6). Расчетная численность потомков в 8 из 10 лет второго периода (19891998 гг.) заметно меньше, чем можно было ожидать исходя из численности родителей. Остатки регрессии 7 в первом периоде в среднем были близки к 0 (табл. 6), во втором периоде их среднее значение, при отсутствии значимых асимметрии и эксцесса распределения, составило -2053,74 ± 922,06 тыс. экз. Остатки двух периодов различаются как по средним значениям (Рт = 0,025), так и по дисперсиям распределений (Рр = 0,018). Таким образом, можно предположить, что либо рождаемость, либо выживаемость потомков во втором периоде в среднем была значительно меньше, чем в первом.
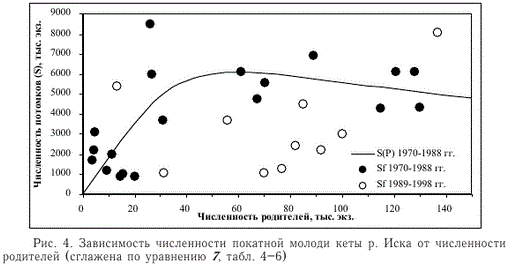
Рис. 4. Зависимость численности покатной молоди кеты р. Иска от численности родителей (сглажена по уравнению 7, табл. 4-6)
Fig. 4. Dependence of the chum juveniles abundance in the Iska river on their parents abundance (smoothed by Equation 7, see Tables 4-6)
Таблица 4
Последовательность построения модели, описывающей зависимость численности покатной молоди кеты р. Иска от численности родителей и метеорологических факторов в 1970-1988 гг.
В уравнение 7 входят следующие параметры: плотностно независимая удельная рождаемость и выживаемость молоди, определяемые региональной спецификой условий воспроизводства (коэффициент а); удельная рождаемость, определяемая степенью удовлетворения производителей в нерестовых площадях (Ь/Р); скорость снижения удельной рождаемости в связи с ростом плотности производителей (коэффициент с). Нет оснований полагать, что во втором периоде уменьшилась площадь нерестилищ кеты или изменилась видовая норма реакции на рост плотности производителей. Одна из возможных причин уменьшения рождаемости или выживаемости может быть связана с ухудшением условий воспроизводства.
Для проверки данной гипотезы необходимо описать связь численности потомков не только с численностью родителей, но и с влиянием метеорологических факторов. Поскольку описание всего материала уравнением 7 невозможно, модель разрабатывали только для первого периода. Принцип ее построения такой же, как и при построении первого варианта (уравнение 6), описание этапов и результаты приведены в табл. 4-6.
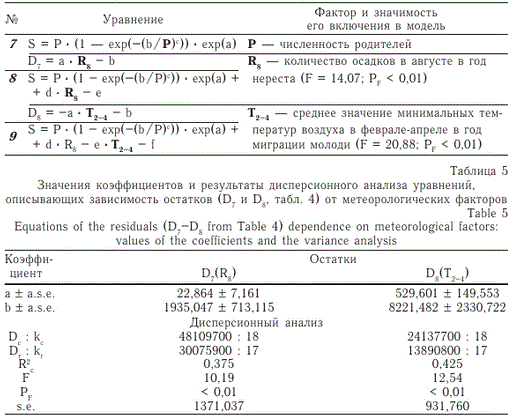
Из результатов построения второго варианта модели (рис. 5) следует, что при самых неблагоприятных погодно-климатических условиях популяция может существовать лишь при численности родителей 30-150 тыс. экз.
При равных условиях (здесь и далее на среднемноголетнем уровне) до 1989 г. численность потомков в среднем должна составлять 5,2 млн экз. При таких же
Таблица 6
Значения коэффициентов и результаты анализа уравнений 7—9 (табл. 4), описывающих зависимость численности покатной молоди кеты р. Иска от численности родителей и метеорологических факторов в 1970-1988 гг.
Примечание. Во всех случаях s.e. асимметрии = 0,524, s.e. эксцесса = 0,766.
Рис. 5. Модельная зависимость (уравнение 9) численности покатной молоди кеты р. Иска при наилучших, средних и наихудших условиях
условиях, но минимальном количестве осадков в августе (R8) численность покатной молоди в среднем должна составлять 3,8 млн экз., при максимальном количестве осадков — 8,4 млн экз. При максимальных значениях температуры воздуха в период с февраля по апрель (Т2-4) и прочих равных условиях численность покатной молоди может составить 2,9 млн экз., при таких же условиях, но минимальных значениях температуры — 7,3 млн экз.
При фиксированной численности родителей отношение максимально возможного количества потомков (при наилучших условиях воспроизводства) к минимально возможному количеству непостоянно, оно зависит от численности родителей. Минимальное отношение данных показателей наблюдается при пропуске на нерест около 87 тыс. рыб (точка максимума) и составляет 3,27 раза.
Судя по значениям критериальной статистики (табл. 6), учитывая отсутствие сериальной корреляции остатков первого порядка ^ = 1,597 > dU = 1,413; Р < 0,01), результат построения модели можно признать удовлетворительным. Дополнение модели метеорологическими факторами увеличило долю объясненной дисперсии зависимой переменной в первом периоде на 39,7 %, но прогноз на второй период по уравнению 9 в 8 из 10 случаев по-прежнему остался завышенным (рис. 6). Остатки составили в среднем -1760,52 ± 934,72 тыс. экз. В 5 из 10 случаев во втором периоде численность потомков оказалась меньше, чем можно было ожидать при наихудших условиях воспроизводства (см. рис. 5).
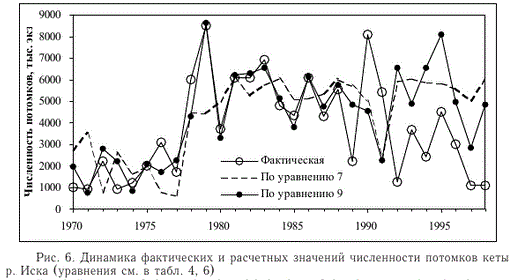
Рис. 6. Динамика фактических и расчетных значений численности потомков кеты р. Иска (уравнения см. в табл. 4, 6)
Попытка подогнать модель ко всем данным оказалась безуспешной даже при фиксированных значениях коэффициентов a-c. Таким образом, в отличие от первого варианта модели (уравнение 6) слишком низкую численность потомков большинства лет после 1988 г. не удалось объяснить влиянием климатических факторов.
Первый вариант модели (уравнение 6), в отличие от второго (уравнение 9), обладает хорошими прогностическими свойствами, поскольку прогноз на второй период, основанный на подгонке модели к данным за первый период, вполне приемлем, хотя при этом пришлось исключить последний фактор (Pg) вследствие больших ошибок коэффициента f (табл. 1-3). При таком тестировании первого варианта модели коэффициент парной корреляции между эмпирической и расчетной численностью потомков во втором периоде равен 0,958, фактические значения и отклик модели не различаются ни по средним значениям (PT = 0,132), ни по дисперсиям (PF = 0,116). Отметим также, что в первом периоде уравнением линейной регрессии S(P) объяснимо 33 % численности потомков, а линейной регрессии S(R8) — лишь 22,3 %, что ставит под сомнение приоритет фактора R8 при разработке первого варианта модели.
Таким образом, учитывая, что относительная среднеквадратическая ошибка уравнения 6 почти на 10 % меньше, чем уравнения 9, можно заключить, что формально по всем параметрам оно имеет явные преимущества перед более обоснованным и “лаконичным” уравнением 9. Единственная причина, которая может реабилитировать уравнение 9, ассоциируется с падением уровня экономики в конце 80-х — начале 90-х гг. прошлого века (карточная система распределения основных продуктов, задержки выплаты заработной платы, развал предприятий и т.п.). Тогда население, имеющее доступ к природным ресурсам, вероятно, перешло на самообеспечение, не ограничиваясь количеством рыбы, необходимым для собственного потребления. Рыбоучетное заграждение на р. Иска установлено в нижнем течении, выше по течению река интенсивно посещается населением с. Власьево и г. Николаевск-на-Амуре. Вполне очевидно, что фактическая численность рыб, размножавшихся в годы экономического кризиса, была значительно меньше, чем было учтено на рыбоучетном заграждении.
Если гипотеза о непостоянстве ошибок измерения основной факториальной переменной (Р) верна, то причины плохого описания данных второго периода уравнением 9 вполне понятны: возросшие ошибки измерения, являющиеся дополнительной причиной разброса эмпирических значений численности потомков относительно линии регрессии (см. рис. 4), нелогично объяснять влиянием климатических факторов. В таком случае необходимо ставить под сомнение качество первого варианта модели, которая хорошо объясняет изменчивость численности потомков как за все время исследования, так и раздельно по периодам. Хорошее описание материала этой моделью при низкой точности и непостоянстве ошибок измерения частично можно объяснить переподгонкой (Хилборн, Уолтерс, 2001; Дрейпер, Смит, 2007) — привлечением к описанию избыточного числа предикторов. По меньшей мере это относится к переменной Р , включение которой увеличивает долю объясненной дисперсии лишь на 1,8 % (см. табл. 3), при этом вероятность значимого улучшения качества модели относительно низка (см. табл. 1). Отметим также, что стандартная процедура определения данной вероятности в пошаговом регрессионном анализе дает завышенные результаты (Дрейпер, Смит, 2007).
Из тихоокеанских лососей в р. Иска кроме кеты размножается горбуша, и, если гипотеза о непостоянстве ошибок измерения численности родителей верна, было бы странным, если бы во втором периоде завышалась только численность производителей кеты. Численность горбуши р. Иска приблизительно на порядок больше, чем кеты, вероятно поэтому браконьерство меньше повлияло на точность оценки численности родителей и весь материал за эти же годы удовлетворительно описывается уравнением 7 (Островский, 2011). Однако, если данные по горбуше также разделить на два периода, как и в случае с кетой (рис. 4), 8 из 10 точек второго периода тоже расположатся ниже линии регрессии (остатки во втором периоде составляют в среднем -4438,5 ± 1888,9). Следовательно, дефицит потомков во втором периоде существовал и у горбуши, причем, как и в случае с кетой, он не был полностью устранен введением в модель метеорологических факторов: ее остатки составили в среднем -1388,2 ± 757,6 тыс. экз.
Таким образом, гипотеза о систематическом завышении оценки численности производителей вследствие браконьерства не лишена оснований, соответственно второй вариант модели, видимо, более реалистичен. К недостаткам второго варианта модели, как и первого, можно отнести то, что в силу аддитивного способа включения в модель погодно-климатических факторов теоретически (хотя экстраполяция в данных случаях лишена смысла) не исключена вероятность получения аномальных значений численности потомков при экстремальных условиях среды (рис. 5).
Этого недостатка можно избежать, если удастся построить мультипликативную модель, аналогичную по структуре модели, разработанной для горбуши данной реки (Островский, 2011). Из наших попыток разработки такой модели для первого периода наиболее успешным оказался вариант, в котором зависимость численности потомков от численности родителей описали уравнением Рикера (1979):
S = P • exp (a - P/b) (10).
Данный вариант интересно сравнить с первыми двумя вариантами, поэтому кратко остановимся на его описании. Уравнение 10 было дополнено влиянием на зависимую переменную количества осадков (R — среднее значение количества осадков в период с июля по ноябрь в год нереста и в апреле следующего года):
S = P • exp (a - P/b + c • R7-11+4) (11).
В полном виде третий вариант модели выглядит следующим образом:
S = P • exp (a - P/b + c • R7-11+4 - d • T2-3) (12),
где Т2-3 — среднее значение минимальных температур воздуха февраля и марта в год миграции молоди. Регрессии, описывающие остатки уравнений 10 и 11 влиянием соответствующих факторов (R и T ), значимы с вероятностью более 99 %, значения коэффициентов данных уравнений в пределах 95 %-ных доверительных интервалов больше 0, коэффициент детерминации в первом случае составил 0,661, во втором — 0,321. Гипотеза об отсутствии существенного улучшения качества модели введением фактора R7-11+4 отвергается с вероятностью более 99 %, фактора T2-3 — более 95 %.
Точность данной модели низка, относительная среднеквадратическая ошибка равна 50,2 %. Если судить по бутстреп-оценкам коэффициентов и их ошибок (табл. 7), можно заключить, что модель (уравнение 12) устойчива к варьированию входных данных. Остатки модели практически равны 0, их распределение близко к нормальному, автокорреляция первого порядка отсутствует (d > 1,685; P < 0,01). С вероятностью более 99 % модель значима, но коэффициент детерминации несколько меньше, чем у первого и второго вариантов. Динамика расчетных частот принципиально не отличается от второго варианта (рис. 6) — оба не пригодны для прогноза на второй период.
Отношение максимальной расчетной численности потомков при наилучших условиях воспроизводства к минимальной численности постоянно и составляет 4,92 при любом значении численности родителей больше 0 (рис. 7). Аналогичные результаты получаются при использовании (в качестве стартового) уравнения 7 вместо уравнения 10, но значение критерия Фишера в таком варианте модели оказывается меньше, чем по уравнению 12, за счет большего числа коэффициентов.
Таким образом, наиболее простая и логичная модель оказывается наименее точной. Вполне очевидно, что выбор одного из трех рассмотренных вариантов модели не может быть основан на формальном использовании критериальной статистики вследствие неопределенности ошибок измерения численности родителей и потомков. Так, например, если ошибка измерения потомков каждый год составляет около 20 %, то первый вариант модели, детерминация которого составляет более 90 %, менее реалистичен, поскольку им объясняется более 10 % дисперсии, определяемой ошибкой измерения. Хотя, с другой стороны, величина ошибки измерения тоже может зависеть от погодных условий, косвенно учтенных в модели. Если вопреки нашим предположениям ошибка измерения численности родителей минимальна и постоянна во времени, то менее реалистичны последние два варианта модели.
Разработка нескольких вариантов модели при имитационном моделировании — обычное явление. Вследствие ограничения рукописи по объему приходит-
Значения коэффициентов и результаты анализа уравнений 10—12, описывающих зависимость численности покатной молоди кеты р. Иска от численности родителей и метеорологических факторов в 1970-1988 гг.
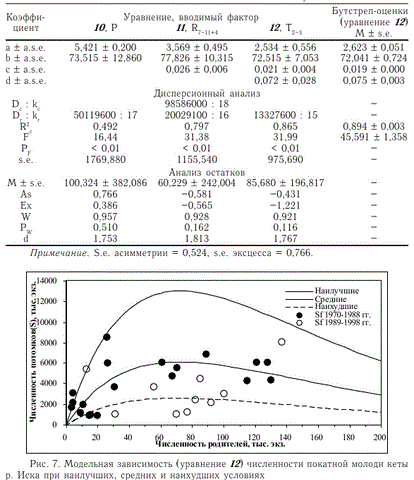
Примечание. S.e. асимметрии = 0,524, s.e. эксцесса = 0,766.
Рис. 7. Модельная зависимость (уравнение 12) численности покатной молоди кеты р. Иска при наилучших, средних и наихудших условиях
ся выбирать “окончательный вариант” для опубликования, что привносит в результаты элемент субъективности и порождает скептическое отношение многих исследователей к моделированию в целом. Но использование данного метода оправдано не только тем, что он позволяет выявлять закономерности в разрозненных явлениях, но и тем, что он предоставляет “общий язык” для сравнения различных популяций и видов. Сравним полученные результаты с результатами аналогичных исследований осенней кеты р. Хор (Островский, Подорожнюк, 2009) и горбуши р. Иска (Островский, 2011). Для упрощения сравнения обозначения факторов из различных источников приведем к системе, принятой в данной работе (табл. 8).
Таблица 8
Уравнения, описывающие зависимость численности молоди кеты и горбуши рек Хор и Иска от численности родителей и метеорологических факторов

Последовательность включения факторов в модель зависит от силы их влияния, первыми включаются факторы, наиболее важные в статистическом плане. Как следует из сравнения трех вариантов описания динамики численности потомков кеты р. Иска (табл. 9), если модели адекватно отражают свойства объекта, то порядок включения основных факторов, направление связи с зависимой переменной и соответственно относительная роль факторов в динамике практически не зависят от архитектуры модели. Качественные расхождения в описании кеты р. Иска разными вариантами модели появляются лишь после включения первых трех факторов (табл. 9), влиянием которых в линейной модели (вариант 1) объяснимо более 70 % дисперсии численности потомков (см. табл. 3).
Таблица 9
Последовательность (1-6) включения факторов в модели и направление их связи (+, -) с численностью покатной молоди

* Направление связи зависит от численности родителей.
Вполне возможно, что дополнения первого варианта модели последними двумя факторами (Т5 и Р10) обусловлены упрощенным описанием нелинейной связи S(P) линейной функцией и случайной корреляцией остатков с данными переменными. Однако положительная связь численности потомков с температурой воздуха в конце весны отмечалась и у кеты р. Хор (табл. 9). Механизм реализации положительной связи зависимой переменной с количеством осадков
в начале зимы в данном варианте модели, по-видимому, объясним снеговой защитой нерестовых бугров от промерзания, т.е. он принципиально не отличим от предполагаемого механизма влияния температуры воздуха в начале зимы на численность кеты р. Хор и горбуши р. Иска (Островский, 2011; Островский, Подо-рожнюк, 2011).
Возможные механизмы влияния остальных факторов на численность потомков рассматривались ранее (Островский, Подорожнюк, 2009; Островский, 2011), поэтому мы не будем возвращаться к их обсуждению, за исключением обсуждения причин положительного влияния на зависимую переменную количества весенних осадков (третий вариант модели). Если данная связь не случайна, тогда, возможно, она обусловлена положительной корреляцией количества весенних осадков с весенним уровнем и скоростью течения воды и, соответственно, с улавливаемостью мальков и их учтенной численностью. По крайней мере факт избегания стандартных ловушек Таранца молодью кеты при низкой скорости течения подтверждается визуальными наблюдениями.
Принципиальное отличие всех вариантов модели, разработанных для кеты р. Иска, от моделей, рассмотренных ранее (табл. 9), состоит в отсутствии связи численности потомков с температурой воздуха летом. Причина данного расхождения очевидна — горбуша р. Иска мигрирует в реку в самое жаркое время сезона, в среднемноголетнем плане 50 % производителей горбуши заходит на нерест в 6-й пентаде июля. Кета заходит в эту реку на месяц позже, кульминация ее нерестового хода приходится на 6-ю пентаду августа, когда температура воздуха и соответственно воды меньше. Еще позже на нерест мигрирует осенняя кета Амура, но температура воды во время ее миграции не меньше, а больше, чем в реках Сахалинского залива, в который впадает р. Иска (Островский, Подорожнюк, 2011).
Заключение
Сравнение трех вариантов описания материала по кете р. Иска позволяет заключить, что при равной численности родителей наиболее многочисленные ее поколения наблюдались в годы обильных осадков во время нерестовой миграции и нереста рыб, особенно если температура воздуха в конце зимы и начале весны была низкой. Влияние данных факторов выявляется в любом из вариантов модели. Не исключено также, что численность молоди на стадии покатной миграции положительно связана с количеством осадков в начале зимы и весной, а также с температурой воздуха в конце весны.
Наиболее общей чертой сравниваемых группировок лососей является тесная связь численности потомков с численностью родителей, а также одинаковое направление связи конкретного метеорологического фактора с численностью потомков любой группировки, для которой такая связь выявлена. Основные причины изменчивости численности поколений кеты сравниваемых рек (и отчасти горбуши) сходны, но роль конкретного фактора (сила влияния) может быть специфичной для каждого вида и места воспроизводства.
Учитывая полученные результаты, можно заключить, что период нерестовой миграции, нереста и раннезимний период инкубации икры являются критическими для формирования урожайности поколений всех рассмотренных группировок тихоокеанских лососей. Кроме того, можно отметить, что численность потомков кеты, в отличие от горбуши, может зависеть от условий, в которых они развиваются в период от выклева до перехода на смешанное питание и, возможно, в период нагула в реке.
Островский Владимир Иванович, кандидат биологических наук, начальник отдела, e-mail: Этот адрес электронной почты защищён от спам-ботов. У вас должен быть включен JavaScript для просмотра..