Зависимость числености потомков летней кеты (ONCORHYNCHUS KETA) Р. Амур от числености родителей
В течение практически всего прошлого века при одинаковом промысловом усилии летней амурской кеты Oncorhynchus keta вылавливалось примерно в 5 раз меньше, чем осенней кеты. Меньшие запасы и вылов летней кеты объяснялись вдвое меньшей абсолютной плодовитостью, вдвое меньшей площадью нерестилищ, большей зависимостью эффективности ее воспроизводства от внешних факторов.
Основываясь на существующих объяснениях причин низкого репродуктивного потенциала летней кеты, невозможно объяснить ее высокие уловы в начале прошлого и текущего веков, когда они превышали уловы осенней кеты. Приведены результаты анализа зависимости численности половозрелых потомков летней кеты от численности родителей, проведенного для выявления причин изменчивости уловов и запаса. При оценке численности родителей и потомков исходили из промысловой статистики и литературных сведений, согласно которым промысел изымает 50 % запаса амурской летней кеты. Связь рассматриваемых переменных аппроксимировали оригинальным уравнением. Согласно полученным результатам, максимальная численность пополнения амурской летней кеты, в среднем близкая к 19,08 млн экз., вполне сопоставима с численностью осенней кеты в урожайные годы. Такое пополнение наблюдается при нересте всего около 1,94 млн экз. В то же время при малой численности родителей скорость роста популяции очень низкая. Эти результаты, на наш взгляд, объясняют причины медленного восстановления запаса летней кеты и потенциальную возможность численного равенства запасов кеты обеих рас. Для поддержания запаса летней кеты на высоком уровне необходим пропуск оптимального количества родителей, но диапазон значений их численности, предполагающий хороший «урожай», очень узкий и пропустить на нерест заданное количество производителей в условиях самой крупной кетовой реки очень сложно.
Введение
Первая попытка описания зависимости численности потомков летней кеты On-corhynchus keta от численности родителей была предпринята на основе материалов 1947-1957 гг., приведенных в работе В.Я. Леванидова (1969), из которых следовало, что максимум численности потомков летней кеты, в среднем близкий к 4 млн экз., наблюдался при нересте около 1 млн рыб, в то время как численность потомков осенней кеты достигала максимума, близкого к 20 млн экз., при нересте 4 млн рыб (Ostrovsky, 2002). Учитывая соотношение запасов летней и осенней кеты, отражаемое в уловах в этот период при практически одинаковом промысловом усилии, данный результат казался вполне правдоподобным.
Меньшие уловы и запас летней кеты по сравнению с осенней объяснялись меньшей индивидуальной абсолютной плодовитостью, менее стабильными условиями нереста и инкубации икры, примерно вдвое меньшим по сравнению с осенней кетой нерестовым фондом (Бирман, 1954, 2004). Однако в годы исторического максимума вылова амурских лососей (1909-1913 гг.) уловы летней кеты превышали уловы осенней, что противоречит представлениям о ее меньшем репродуктивном потенциале. В этот период ее среднегодовой вылов составлял около 36 тыс. т (55 % суммарного улова амурской кеты), т.е. в 4 раза больше, чем следовало из выявленной связи «запас-пополнение» (Ostrovsky, 2002). Данное противоречие отчасти снималось всеобщим скептическим отношением к промысловой статистике начальных лет мониторинга. Скепсис основывался на почти вековом наблюдении относительно низких уловов летней кеты после 1913 г. С 1914 г. до конца прошлого века в среднем за год ее вылавливали около 1,8 тыс. т, в то время как осенней кеты — 9,7 тыс. т.
С начала XXI в. уловы кеты обеих рас вновь начали увеличиваться, достигнув в 2014 г. 36 тыс. т, причем в некоторые годы, как и в начале прошлого века, в уловах вновь доминировала летняя кета. В 2012 г. ее выловили более 8 млн экз. (17 из 28 тыс. т амурской кеты). Следовательно, запас в данном году, как и в начале прошлого века, в несколько раз превышал теоретический максимум, рассчитанный по описанной зависимости «запас-пополнение» (Ostrovsky, 2002), что подтверждает ее несостоятельность. Таким образом, несмотря на более чем вековую историю изучения амурской летней кеты феномен ее высоких уловов, сопоставимых с уловами осенней кеты, при наличии явных признаков меньшего репродуктивного потенциала пока не имеет непротиворечивого объяснения.
Цель данной работы — анализ связи численности потомков летней кеты с численностью родителей. Знание связи данных переменных необходимо не только для разрешения несоответствия представлений о репродуктивных возможностях летней и осенней кеты их вылову, но и для разработки прогноза и рационального использования запаса.
Материалы и методы
Работа основана на опубликованных данных по вылову амурской кеты в период 1908-1986 гг. (Яновская и др., 1989) и более поздних данных по вылову, ежегодно предоставляемых организациями, контролирующими вылов (рис. 1). Промысловая статистика амурской кеты включает уловы в районе малых рек южной части амурского лимана. Промысел в этом районе затруднен ввиду отдаленности от центров переработки рыбы, мелководности прибрежий, затрудняющих передвижение плавучих средств, особенно при обычном для лимана сильном ветре, поэтому вылов здесь мал и практически не влияет на динамику вылова амурских лососей.
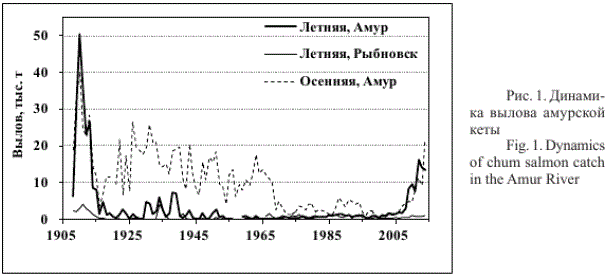
Рис. 1. Динамика вылова амурской кеты
Многие исследователи считают, что в русле и лимане Амура с относительно постоянной интенсивностью облавливается около 50 % запаса амурских лососей (Смирнов, 1947; Леванидов, 1964, 1969; Рослый, 2002). Кроме того, амурская летняя кета вылавливается как вынужденный прилов при промысле горбуши в районе пос. Рыбновск на северо-западном побережье о. Сахалин. Учитывая данные сведения, в качестве рабочей гипотезы можно принять, что численность прошедших на нерест амурских лососей равна вылову в русле и лимане Амура, а запас (численность или биомасса рыб, вернувшихся из моря в данном году) — удвоенному вылову в данных районах в сумме с выловом в районе пос. Рыбновск.
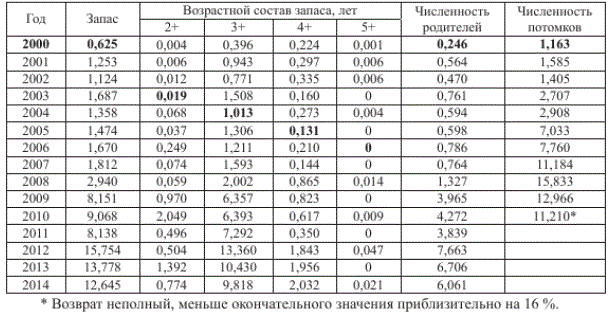
Закономерности воспроизводства анализировали на основе материалов по вылову в 2000-2014 гг. Численность запаса и выловленных рыб рассчитывали делением биомассы на среднюю массу тела одной особи летней кеты в данном году. Зная численность запаса и возрастной состав, рассчитали абсолютную численность рыб в возрастных группах (табл. 1). Суммируя численность рыб, принадлежащих к одному поколению (для примера, в табл. 1 выделены значения для поколения 2000 г.), рассчитали численность потомков.
Материал анализировали методами регрессионного и дисперсионного анализов, изложенными в справочной литературе (Поллард, 1982; Дрейпер, Смит, 2007; Медик, Токмачев, 2007). В работе приняты следующие обозначения: Рр Рт — уровни значимости соответствующих критериев Фишера и Стьюдента; индексы f и st — фактические и табличные значения критериев; s.e., а^.е. — стандартная и асимптотическая стандартная ошибки; Dг, Dc — соответствующие индексам остаточная и общая скорректированная сумма квадратов остатков; к, кс — соответствующие индексам числа степеней свободы; R2c — скорректированный коэффициент детерминации; М — среднее арифметическое значение; As, Ех — коэффициенты соответственно асимметрии и эксцесса. Строчными буквами в уравнениях обозначены коэффициенты, значения которых подбирали методом итераций, реализованным в пакете прикладных программ SYSTAT. Численность рыб приведена в миллионах экземпляров.
Таблица 1
Исходные данные для расчета численности родителей и потомков летней кеты р. Амур, млн экз.
где R — численность потомков, Р — численность родителей, а-с — коэффициенты, значения которых приведены в табл. 2 (вариант а). Для сравнения на рис. 1 приведена кривая воспроизводства осенней амурской кеты, построенная по литературным данным (Островский и др., 2015). Уравнение (1) ранее использовалось для описания зависимости численности покатной молоди горбуши охотского района от численности родителей (Островский, Пономарев, 2009). Его выбор в данной работе основан, с одной стороны, на внешнем сходстве графиков, с другой — на лучших его статистических показателях качества аппроксимации по сравнению с иными наиболее часто используемыми для этих целей уравнениями.
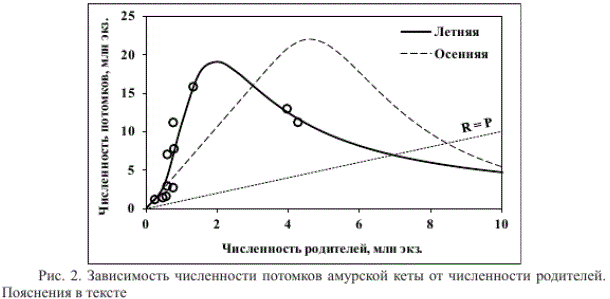
Рис. 2. Зависимость численности потомков амурской кеты от численности родителей. Пояснения в тексте
Таблица 2
Значения коэффициентов и результаты дисперсионного анализа уравнения, описывающего зависимость численности потомков амурской летней кеты от численности родителей
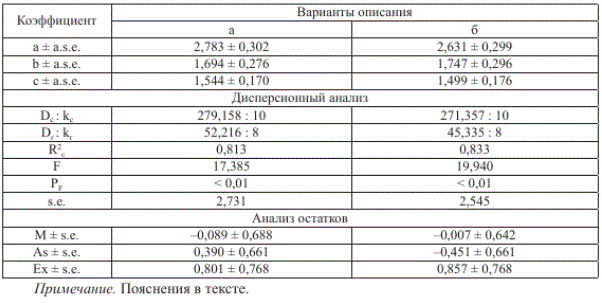
Гипотеза о равенстве коэффициентов уравнения нулю (проверена по Б-критерию Фишера, табл. 2, вариант а) отвергается с вероятностью более 99 %. Средняя численность потомков, рассчитанная по уравнению (1) (6,975 ± 1,394 млн экз.), практически не отличается (Т = 0,042 < Т = 2,086, Рт > 0,05) от фактического значения данного показателя (6,887 ± 1,593 млн экз.). Не различаются и дисперсии распределений сравниваемых величин (Б(. = 1,307 < Б = 3,179, Рр > 0,05). Отсутствие значимых асимметрии и эксцесса распределения остатков регрессии (табл. 2, вариант а) свидетельствует о том, что оно близко к нормальному.
Приведенные результаты позволяют заключить, что уравнение адекватно аппроксимирует реальную изменчивость численности потомков летней амурской кеты в описываемом периоде, однако вопрос об устойчивости уравнения, а следовательно и о его прогностической ценности, остается открытым.
Для проверки устойчивости уравнения часто используется процедура разделения временного ряда на два интервала, первый предназначен для построения и оценки параметров модели, второй для проверки ее прогностических свойств. В нашем случае такая процедура неприемлема, поскольку временной ряд слишком короток. Подобрать уравнение для временного ряда большей длительности по используемой нами схеме непросто в силу того, что данные по возрастному составу и среднегодовой массе тела за предшествующие годы разобщены в различных источниках, неполны и не всегда совпадают.
По этой причине мы попытались упростить схему расчетов. Численность родителей и потомков вычислили делением их биомасс на среднемноголетнюю массу тела (2,208 кг), возраст созревания всех потомков приняли равным 3+ лет (в действительности в данном возрасте — около 78 % рыб), коэффициент эксплуатации равным 50 %. В соответствии с принятыми допущениями численность родителей равна численности рыб, выловленных в Амуре и лимане, а численность потомков — удвоенной численности рыб, выловленных в данном районе, в сумме с уловом в пос. Рыбновск через 4 года после нереста.
Такой вариант использования материала, при его аппроксимации уравнением (1), практически не повлиял на качество описания (табл. 2, вариант б), о чем свидетельствуют близкие значения коэффициентов уравнений обоих вариантов (а и б). Эмпирические
и рассчитанные по второму варианту значения переменных в 2000-2014 гг. довольно близки, о чем свидетельствуют малые значения стандартизированных остатков (рис. 3), размерные остатки практически нормально распределены относительно 0, скорректированный коэффициент детерминации составляет 0,833 (табл. 2, вариант б). Остальные статические параметры также свидетельствуют о том, что результаты анализа первичных данных в таком варианте не уступают результатам их использования по варианту а.
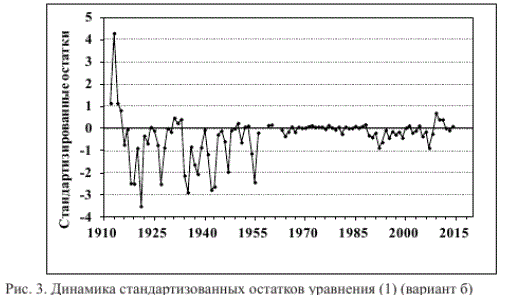
Рис. 3. Динамика стандартизованных остатков уравнения (1) (вариант б)
Таким образом, намеренное огрубление расчетов не привело к существенным потерям точности описания данных по вылову амурской кеты в зоне интерполяции, позволяя при этом составить ретроспективный «прогноз» и оценить устойчивость уравнения. «Прогноз» численности потомков по уравнению (1) (вариант б) на период 1999-1955 гг. оказался вполне удовлетворительным (рис. 3).
Некоторое завышение «прогноза» численности потомков (по сути «прогноза» вылова) в конце 1980-х — начале 1990-х гг. (рис. 3) с использованием любого из вариантов расчета, вероятно, объяснимо неполной отчетностью о вылове лососей в годы экономического кризиса. Аналогичные для этих лет последствия, отражающиеся на соотношении «родители-потомки», выявлены при исследовании горбуши и кеты р. Иска (Островский, 2012). Занижение данных по уловам амурских лососей рыбаками наблюдалось и в начальные годы текущего века (Капланова и др., 2004). Возможные причины неадекватного отклика модели в первой половине прошлого века рассмотрим ниже.
Учитывая сходство результатов описания связи «запас-пополнение» в обоих вариантах обработки материала в зоне интерполяции и относительную устойчивость уравнения в варианте б, можно допустить, что более детализированный вариант а также устойчив и при сохранении условий воспроизводства прогноз на основе данного варианта будет приемлем и на ближайшее будущее.
Второй вариант расчета (б) основан на сведениях по биомассе улова, трансформированной стандартной процедурой в численность родителей и потомков, поэтому можно заключить, что биомасса улова в текущем году должна зависеть от биомассы улова 4 года назад и влиять на биомассу улова, который будет наблюдаться через 4 года. Такой анализ исходных данных избавляет от необходимости принятия рабочей гипотезы относительно степени эксплуатации и может быть использован для прогноза величины улова летней кеты при постоянном промысловом усилии, однако он не отвечает целям работы, поскольку не позволяет сопоставить кривые воспроизводства летней и осенней кеты (см. рис. 2).
Таким образом, полагаем, что уравнение (1) удовлетворительно описывает современную динамику численности амурской летней кеты и приемлемо для разработ-
ки предварительных прогнозов запаса. Кратко рассмотрим его свойства. Согласно полученным результатам (вариант а), максимальная численность потомков амурской летней кеты, в среднем равная 19,083 млн экз., наблюдается при нересте 1,937 млн рыб. Максимальная разность численности потомков и численности родителей, составляющая в среднем 17,189 млн экз., наблюдается при нересте 1,855 млн рыб (оптимум пропуска). При нересте более 6,931 млн экз. кеты численность потомков в среднем меньше численности родителей.
Поскольку численность родителей 2009-2014 гг. известна (см. табл. 1), по уравнению (1) (вариант а) можно рассчитать численность потомков (Rt) и запаса до 2018 г. (табл. 3). В связи с неодновременным созреванием кеты потомки, например, от нереста в 2009 г. (в табл. 3 выделены) вернутся из моря в 2012-2015 гг., их возрастной состав и масса тела приняты равными среднемноголетним значениям соответствующих переменных летней кеты.
Таблица 3
Прогноз численности и биомассы запаса летней кеты р. Амур
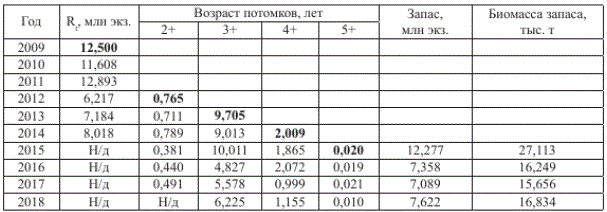
Согласно расчетам, по принятой схеме в 2015 г. ожидается возврат 12,277 млн особей (27,113 тыс. т) амурской летней кеты (табл. 3). Необходимо отметить, что расчетная численность потомков соответствует среднестатистическому ожиданию реализации прогноза при постоянстве условий воспроизводства. В то же время известно, что повышенная температура воды в Амуре в 2011 г. (до 24 оС в конце второй и до 28 оС в конце третьей декад июля), сопровождающаяся массовой донерестовой гибелью половозрелых рыб, являясь мощным стрессором, оказывает не только прямое влияние на количество потомков, но и косвенное, снижая их выживаемость (Подлесных, Пащенко, 1997; Подлесных, 2000).
Если промысловая статистика первой половины ХХ века верна, тогда неадекватный отклик модели в период 1908-1955 гг. (рис. 3) свидетельствует о том, что либо уравнение (1) не идеально, либо градации факторов среды часто выходили за пределы адаптационных возможностей рыб, либо принятое допущение о постоянном изъятии 50 % запаса не всегда было близко к действительности. Зависимость численности потомков от численности родителей в этом периоде (рис. 4) действительно может быть более сложной по сравнению с зависимостью, представленной на рис. 2. График данной зависимости двухвершинный, расположение точек в пределах левого пика (рис. 4) весьма сходно с расположением точек, приведенным на рис. 2. Это позволяет предположить, что уравнение (1) можно рассматривать как частный случай более сложной двуцикловой модели (Островский, Семенченко, 2002; Островский, Пономарев, 2009).
С другой стороны, уверенности в более сложном характере зависимости численности потомков от численности родителей нет: все случаи с расчетной численностью родителей более 5 млн экз. (рис. 4) относятся к периоду неконтролируемого лова в 1909-1913 гг., за которым последовало катастрофическое снижение запасов (Навозов-Лавров, 1927; Смирнов, 1947; Бирман, 1954; Никольский, 1954). Вполне вероятно, что
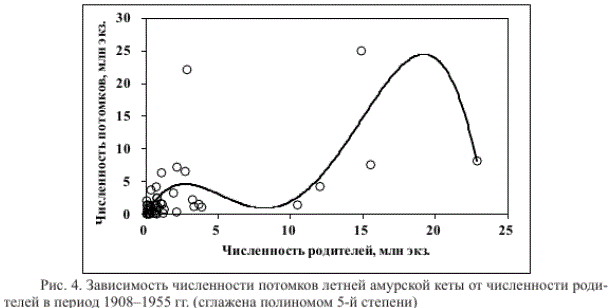
Рис. 4. Зависимость численности потомков летней амурской кеты от численности родителей в период 1908-1955 гг. (сглажена полиномом 5-й степени)
в этот период вылавливали более 50 % запаса лососей, т.е. численность родителей была меньше численности выловленных рыб, соответственно запас был меньше удвоенного вылова. В таком случае все точки для P > 5 млн экз. на рис. 4 должны располагаться левее и ниже, возможно, попадая в область значений левого пика или значений, приведенных на рис. 2.
Кроме того, известно, что в 1925-1930 гг. вводили запрет на промысел летней кеты (Бирман, 1954), соответственно в период 1921-1934 гг. на основе принятой нами гипотезы неверно оценена численность родителей и потомков. Возможно, предположение об изъятии 50 % запаса слишком отличается от реальной промысловой нагрузки и в годы Великой Отечественной войны. Меньшую, чем можно было ожидать, численность потомков летней кеты в первой половине прошлого века (см. рис. 3) некоторые исследователи связывали и с чрезвычайно суровыми зимами 1911-1914 и 1938-1939 гг. (Смирнов, 1947; Бирман, 1954; Леванидов, 1964; и др.).
Таким образом, вполне вероятно, что большие расхождения расчетных и фактических значений улова летней кеты в первой половине прошлого века (рис. 3) связаны с несостоятельностью рабочей гипотезы о постоянном вылове половины запаса в данный период, а также с неучтенной в модели нестабильностью условий воспроизводства.
Если куполообразная форма кривой воспроизводства лососей (см. рис. 2) является следствием убывающей рождаемости по мере роста плотности производителей на нерестилищах (Островский, Семенченко, 2002), то положение точки максимума кривой должно определяться нерестовым фондом. Судя по соотношению площадей нерестилищ осенней и летней кеты (приблизительно 2 : 1), точка максимума кривой воспроизводства осенней кеты должна быть примерно вдвое больше точки максимума летней кеты, что не противоречит полученным результатам (рис. 2). Поскольку форма кривой пополнения зависит от соотношения численностей родителей и потомков, рассчитанных на основе рабочей гипотезы о стабильном изъятии 50 % запаса, соответствие пропорций нерестового фонда пропорциям точек максимума кривых воспроизводства также свидетельствует о достаточной обоснованности рабочей гипотезы.
В отличие от классических моделей «запас-пополнение» типа Бивертона-Холта или Рикера (Рикер, 1979), численность потомков летней кеты, приходящихся на 1 родителя, по нашей модели максимальна не при минимальном значении численности родителей, а при нересте около 1,305 млн рыб. Меньшее соотношение R/P при меньших значениях численности родителей (рис. 2, вогнутая часть восходящей ветви кривой) может быть обусловлено пониженной рождаемостью, например при сверхнизкой плотности производителей отдельных субпопуляций, или повышенной долей рыб, травмированных хищниками.
Такой же эффект может наблюдаться вследствие повышенной смертности потомков при постоянном неучтенном изъятии части запаса независимо от его численности, например при элиминации хищниками. Учитывая, что данное явление не обнаруживается у осенней кеты, полагаем, что оно обусловлено преимущественно постоянно не полностью учитываемым приловом летней кеты при промысле горбуши на о. Сахалин и на амурских промыслах.
Пониженная скорость роста численности летней кеты при малой численности родителей наряду с большей зависимостью ее воспроизводства от климатических факторов (Смирнов, 1975; Бирман, 2004) препятствуют быстрому выходу из депрессивного состояния. В то же время при средних значениях численности родителей скорость роста численности потомков больше, чем у осенней кеты (рис. 2). По этой причине, несмотря на значительно меньший нерестовый фонд, летняя кета очень быстро способна достичь уровня численности, близкого к уровню осенней кеты, что наблюдалось в конце прошлого и начале текущего века (см. рис. 1, 2).
Традиционно 50 % прогнозного значения запаса амурских лососей рекомендуется к вылову. Согласно предварительному прогнозу на основе оценки численности покатной молоди, возможный вылов летней амурской кеты в 2015 г., включая вылов на сахалинских промыслах, рекомендован в объеме 5,965 млн экз. (12,996 тыс. т). Как следует из выявленных нами закономерностей воспроизводства (уравнение (1), вариант а), численность рыб, зарезервированных для нереста (5,965 млн экз.), в среднестатистическом плане обеспечивает возврат 8,158 млн экз. потомков. Однако вылов в 2015 г. и «урожай» потомков можно увеличить, если для нереста оставить 1,855 млн рыб, численность потомков при пропуске на нерест такого количества рыб в среднем должна быть значительно больше (около 19 млн экз.), чем при использовании традиционной схемы.
Рассчитать значение численности потомков (И^ от нереста в 2015-2018 гг. пока невозможно, поскольку неизвестно, сколько рыб отнерестится в эти годы. Однако используя прогнозное значение запаса (табл. 3) и сделав определенные допущения относительно промысловой нагрузки, можно рассчитать численность рыб, оставшихся для воспроизводства, и численность их потомков. Аналогичные расчеты можно продлить в будущее, анализируя наиболее вероятный отклик запаса на управленческие решения.
Так, при отсутствии промысла (к = 0) и внешних воздействий запас модельной популяции неограниченно долго варьирует близ линии устойчивого равновесия (Я=Р = 6,931 млн экз.) — 4 года значений R > Р сменяются 4 годами значений R < Р (рис. 5). При постоянном изъятии 50 % запаса его продуктивность увеличивается (рис. 6), суммарный вылов за 15 лет промысла составляет 75 млн экз. (рис. 7).
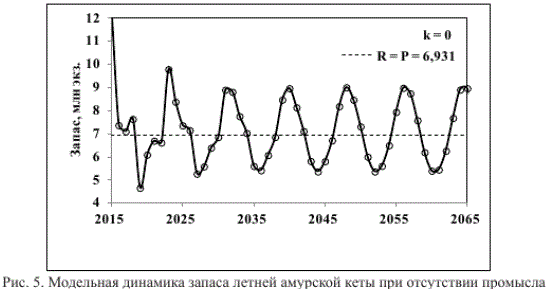
Рис. 5. Модельная динамика запаса летней амурской кеты при отсутствии промысла Fig. 5. Modeled dynamics of summer chum stock in the Amur River without harvesting
Fi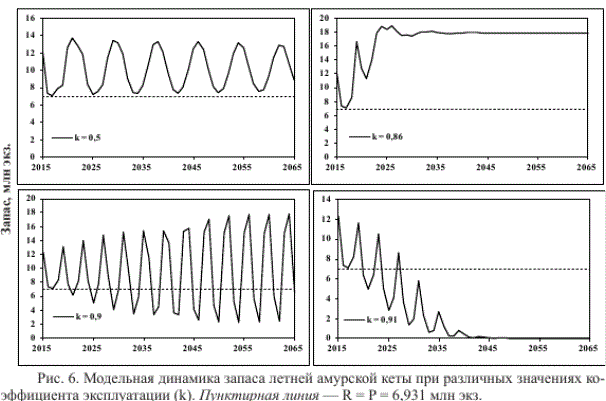
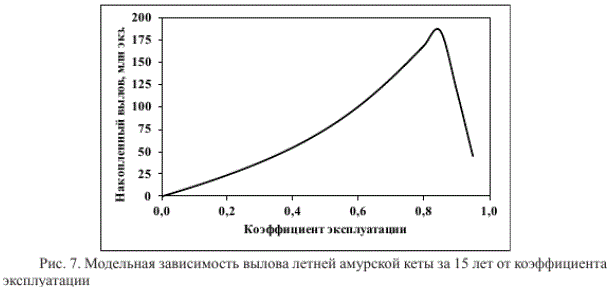
Рис. 7. Модельная зависимость вылова летней амурской кеты за 15 лет от коэффициента эксплуатации
При постоянной промысловой нагрузке 86 % продуктивность запаса больше, вылов с такой интенсивностью в течение 15 лет достигает максимального значения — 186 млн экз. (рис. 7). Если же постоянно изымать 90 % запаса, вылов уменьшается до 119 млн экз., амплитуда его колебаний вновь увеличивается, но, по сравнению с вариантом изъятия 50 % запаса, уменьшается их период. Со временем два года высокой урожайности сменяют два года низкой урожайности. Даже при столь интенсивной эксплуатации продуктивность неурожайных поколений сохраняет способность к восстановлению, но лишь через 80 лет запас обеих неурожайных линий превысит равновесное значение.
Полученные результаты свидетельствуют о том, что причина квазициклических изменений численности популяции не всегда обусловлена циклическими изменениями климата.
При фиксированной интенсивности промысла более 90 % запас деградирует. Если же постоянно пропускать на нерест оптимальное количество родителей (1,855 млн экз.),
вылов за 15 лет составит 214 млн экз., т.е. будет практически равен вылову за такой же период осенней амурской кеты (213-218 млн экз.) при использовании стратегии постоянного остатка (Островский и др., 2015).
Равновесное значение постоянного коэффициента эксплуатации (ке = ^ - Р)/К.), систематическое превышение которого ведет к деградации запаса, зависит от уровня численности родителей. При их низкой численности данное значение меньше, чем при средней (рис. 8), вследствие пониженной скорости роста популяции. Следовательно, если резкое снижение запаса в начале прошлого века могло наступить при изъятии более 90 % запаса (или меньшего изъятия при экстремально неблагоприятных условиях воспроизводства), то для сохранения состояния длительной депрессии достаточно было вылова с меньшей интенсивностью.
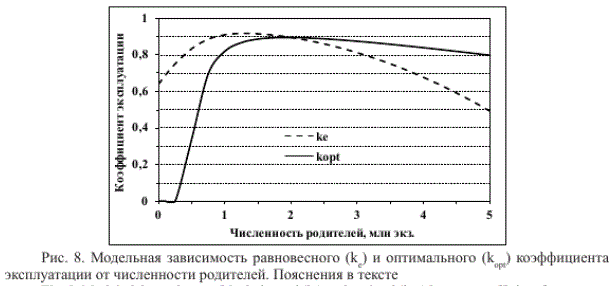
Рис. 8. Модельная зависимость равновесного (ke) и оптимального (k ) коэффициента эксплуатации от численности родителей. Пояснения в тексте
Согласно наилучшей для лососевых промыслов стратегии постоянного остатка, оптимален коэффициент эксплуатации (к ), равный ^ - Р)/Я, где Рор4 — оптимальная численность родителей. При численности родителей менее 0,25 млн экз. R < Р поэтому для скорейшего восстановления запаса целесообразен запрет промысла (к = 0, рис. 8). Однако по причине одновременной миграции летней кеты и горбуши амурского и сахалинского происхождения данное решение трудноосуществимо и должно приниматься с учетом экономической и социальной целесообразности.
Заключение
Таким образом, ни меньшая плодовитость, ни большая зависимость рождаемости от условий воспроизводства, ни меньший нерестовый фонд летней амурской кеты по сравнению с осенней не препятствуют достижению значительного запаса, сопоставимого с запасом осенней кеты. Причина этого — большая скорость роста численности потомков и, соответственно, запаса при средних значениях численности родителей. Из этого следует, что суждение о потенциальной величине запаса по нерестовому фонду, а также экстраполяция учтенной численности рыб в одной реке на остальные реки в пропорции площадей их нерестилищ не всегда корректны.
Не исключая вероятности неблагоприятных условий воспроизводства в прошлом веке, полагаем, что одна из причин длительного периода депрессивного состояния, по-видимому, связана с пониженной скоростью роста численности потомков при малой численности производителей, увеличению последней препятствовал вынужденный прилов летней кеты при промысле горбуши.
Длительность сохранения запаса летней кеты на высоком уровне, при прочих равных условиях, зависит от того, насколько технически удастся пропускать на нерест оптимальное количество производителей, близкое к 1,855 млн экз.
В.И. Островский, О.В. Вершинина, А.П. Шмигирилов*
Хабаровский филиал Тихоокеанского научно-исследовательского рыбохозяйственного центра, 680028, г. Хабаровск, Амурский бульвар, 13а