ВАЛЬЧАКИ АТЛАНТИЧЕСКОГО ЛОСОСЯ, SALMO SALAR L., В ПЕРИОД КАТАДРОМНОЙ МИГРАЦИИ ИЗ РЕК БАЛТИЙСКОГО И БЕЛОГО МОРЕЙ
Часть особей атлантического лосося выживает после первого созревания, мигрирует из
рек к местам нагула на стадии «вальчака», а затем вновь созревает. Доля
повторносозревающих рыб («остатка») в составе нерестовой части разных популяций вида, в
зависимости от естественной и промысловой смертности, может составлять от 1 до 30 % и более [1,2].
Пребывание производителей лосося в реках сопряжено с голоданием,
использованием энергетических ресурсов организма, поэтому отражается на степени
истощения (упитанности), индексах внутренних органов и других особенностях вальчаков.
Длительность периода от захода до катадромной миграции лосося зависит от
географического положения и природно-климатических особенностей водоемов. В реки
южной части ареала он заходит, как правило, позднее, а покидает их раньше, чем на Севере.
Нами изучены в сравнительном аспекте вальчаки лосося из р. Невы бассейна
Балтийского моря и р. Поной бассейна Белого моря (табл. 1). Анализировали также данные
по вальчакам из других рек Кольского п-ова - Варзуги и Лувеньги.
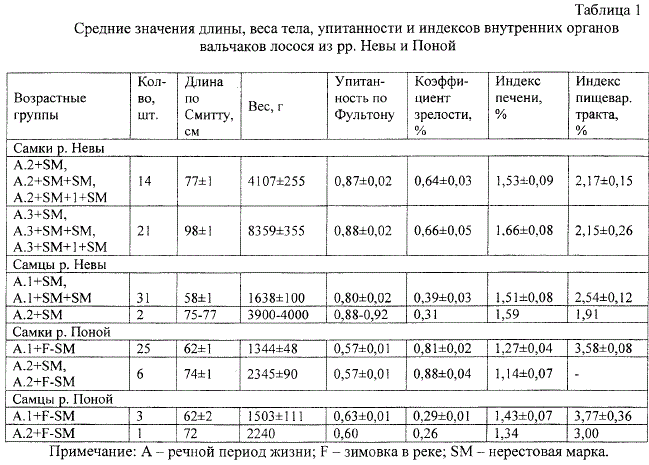
Примечание: А - речной период жизни; F - зимовка в реке; SM - нерестовая марка.
В Неву производители лосося заходят с мая по первую половину октября, созревают во
второй половине октября - начале ноября того же года, а мигрируют из этой реки как
вальчаки в марте - апреле следующего года и появляются в Невской губе Финского залива в
мае, на 15-20 дней раньше, чем смолты. Близкие сроки миграции вальчаков известны в реках
Польши [3]. Таким образом, обычная продолжительность пребывания производителей
лосося в р. Неве составляет 8 - 1 2 месяцев. Лишь отдельные ослабленные самцы могут
выноситься течением в предъустьевое пространство реки еще в конце ноября - декабре.
Невские вальчаки представлены особями обоего пола. Тело у них прогонистое, спина темная,
бока тускло-серебристые с х-образными черными пятнами, брюхо белое, грудные плавники с
черноватыми краями, а брюшные и анальный - сероватые. Чешуя держится прочно, прирост
отсутствует. Вальчаки возобновляют питание в р. Неве уже при температуре воды 0,2-2,0 °С;
в их желудках встречаются рыбы, лягушки, личинки ручейников. У вальчаков из Невской
губы выявлены только переваренные остатки пищи и, в одном случае, искусственная
приманка - «твистер». Желчный пузырь заполнен темно-зеленой желчью. Печень бордового
цвета. Висцеральный жир отсутствует. На пилорических придатках много нематод и
ленточных червей. Яичники розовато-красноватые, с белесовато-сизой собственной
оболочкой, различимой сетью расширенных кровеносных сосудов и широко раскрытой
продольной «щелью». Часто обнаруживаются остаточные икринки: полупрозрачные,
бесцветные или желтовато-оранжевого цвета с жировыми каплями, иногда теряющие тургор.
Количество их может исчисляться сотнями и тысячами штук, а суммарный вес достигает 79
- 268 г и более. Известны случаи поимки самок-вальчаков, у которых полость тела была
заполнена овулировавшей икрой. Яйценосные пластинки лежат рыхло. В них видны
запустевшие фолликулы и ооциты очередной генерации - непрозрачные, беловатого или
светло-желтоватого цвета. Наибольшие значения диаметра их у разных самок - вальчаков,
даже одного возраста, могут составлять от 0,6-0,7 до 1,2-1,3 мм. Для сравнения отметим, что
у ходовых самок в мае ооциты крупнее (2,0 - 2,2 мм), темно-оранжевого цвета и содержат
значительное количество желтка. Семенники розового цвета, лентовидно-уплощенные,
заостренные в краниальной части. Семяпроводы расширенные. Пятна остаточной спермы
встречаются в марте-апреле, но отсутствуют в мае. На гистологических препаратах отмечены
«светлые» сперматогонии типа А, предшествующие началу новой волны сперматогенеза.
Мелкие самцы-вальчаки (A.1+SM) ко времени миграции в Невскую губу обычно теряют
крюк на нижней челюсти, что позволяет им возобновлять питание и созревать повторно. У
крупных же самцов брачные изменения скелета головы необратимы и такие особи не
выживают после завершения сезона нереста. Средние значения коэффициента упитанности у
самок - вальчаков в возрасте A.2+SM; A.3+SM и самцов-вальчаков A.1+SM (см. табл.)
достоверно ниже (Р<0, 01), чем у производителей соответствующих возрастных групп и пола
(А.2+; А.З+ и А.1+) в преднерестовый сезон (1, 24 ± 0,02; 1, 36 ± 0, 03 и 1, 05 ± 0, 03) и ко
времени его завершения, то есть после освобождения от гамет (1,03 ± 0,01; 1,08 ± 0, 04 и 1
,04 ± 0, 02). Средние потери веса самками лосося за период зимовки в р. Неве составляют
около 10 %, а самцами - 15 %.
В бассейне Белого моря значительную часть контингента производителей этого вида
составляют особи осеннего хода, мигрирующие в реки за год до завершения созревания и
нереста. Размножение лосося в условиях Кольского полуострова приходится на сентябрь, а
катадромная миграция большинства вальчаков - на июнь следующего года. Некоторые
ослабленные, погибающие самцы, как и в бассейне Балтики, скатываются поздней осенью в
год созревания. Они характеризуются хорошо выраженным брачным нарядом, а также,
часто, наличием ран и очагов сапролегнии на теле. Сезон массовой миграции вальчаков в
реках Белого моря отчасти перекрывается со временем миграции смолтов. Вальчаки здесь,
как и в бассейне Балтики, при катадромной миграции имеют серебристую окраску [4]. Они
возобновляют питание мелкими рыбами и дрифтом. Обычная продолжительность периода
пребывания в реках непитающихся производителей лосося осеннего хода составляет 22-23
месяца. За это время самки, вернувшиеся в возрасте А.1+ и А.2+, теряют около 49 % веса
тела, а самцы тех же возрастных групп - 53 и 67 %. Вероятно столь значительные потери
ресурсов организма снижают выживаемость и объясняют редкость самцов среди вальчаков
лосося на Севере. В период катадромной миграции они еще не теряют крюк. Средние
значения длины тела по Смитту у вальчаков обоего пола в исследованных нами выборках
больше, чем у заходящих в реки производителей тех же возрастных групп. Аналогичные
различия в размерах и повышенная смертность самцов отмечены финскими и норвежскими
исследователями при исследовании лосося р. Тено [5]. Не исключено, что особи,
вернувшиеся в реки после завершения нагула, продолжают расти, используя резервы
организма, как это происходит у смолтов, содержащихся в экспериментах без пищи.
Коэффициент упитанности у вальчаков из рек бассейна Белого моря достоверно ниже
(Р<0, 01), чем у вальчаков из р. Невы в бассейне Балтики, хотя при заходе из моря лосось
осеннего хода на Кольском п-ове характеризуется более высокими средними значениями
этого показателя: 1,33 ± 0,002 и 1,30 ± 0,003 у самок в возрасте А.1+ и А.2+, а у самцов тех
же возрастных групп - 1,33 ± 0,003 и 1,31 ± 0,003. Достоверные региональные различия
найдены также в значениях коэффициента зрелости, индексов печени и пищеварительного
тракта (табл. 1). Диапазон диаметра наиболее крупных ооцитов очередной генерации у
самок-вальчаков из рек бассейна Белого моря почти такой же, как в р. Неве - 1,0 - 1,2 мм.
Количество же остаточной икры в полости тела значительно меньше, чем у вальчаков из р.
Невы, и обычно исчисляется единицами или несколькими десятками штук. По-видимому,
это объясняется более эффективным участием производителей лосося в естественном
нересте на Кольском п-ове.
И. Г. Мурза, О. Л. Христофоров
Санкт - Петербургский государственный университет, Санкт - Петербург,
bigfish@OC4414. spb. edu