Преднерестовые миграции кеты
Работы по мониторингу морского периода жизни тихоокеанских лососей имеют большое значение в изучении выживаемости поколений, распределения лососей на местах нагула, оценке сроков и динамики преднерестовой миграции, оперативной корректировке прогнозов подходов и управлении береговым промыслом. Особую актуальность представляют исследования, проводимые на конечной фазе морского периода жизнилососей — во время возврата созревающих производителей (рекрутов) к родным водоемам. Проведение судовых экспедиционных работ позволяет заблаговременно (до 2—3 недель) оценить интенсивность подходов производителей и скорректировать сроки начала путины, а также получить данные о распределении миграционных потоков и биологических характеристиках лососей. Накопление рядов лет наблюдений по срокам, условиями интенсивности преднерестовых миграций тихоокеанских лососей дает возможность существенно повысить точность оперативных прогнозов и обеспечивает рациональное управление их запасами. Методической основой оценки уровня и численности подходов лососей с применением лова пелагическими сетями стало предположение о пропорциональности уловов на усилие (улов на 1 сеть длиной 50 м) на путях миграции и величин запасалососей. Многолетние рядыданных уловов на усилие (экз./сеть) позволяют достоверно оценивать связи уловов на усилие с численностью возвратов. Так, например, степень связи дрифтерных уловов горбуши с численностью ее подходов к восточному побережью Камчатки довольно высока (значения R* составляли 0,74—0,87) (Ерохин, 2007а).
Таблица 19. Уловы кеты на единицу усилия (экз./сеть) и ее доля в уловах в разных районах исследований (%) и ее доля в уловах в разных районах исследований (%)")
Проведение исследований морского периода жизнилососей с использованием контрольного дрифтерноголова является важным и необходимым инструментом анализа преданадромных потоков и оценки величин их подходов в регионы Дальнего Востока. Наличие таких данных позволяет осуществлять заблаговременную корректировку уровня, сроков подходов лососей и оперативное управление береговым промыслом (Синяков,
2008).
По результатам проведения в 2001—2003 гг. экспедиционных научно-исследовательских работ получены сведения о сроках, динамике, относительной численности, уловам на усилие и других показателях кеты во время ее преднерестовых миграций в северо-западной части Тихого океана и Охотском морена
этапе продвижения из местнагула к местам воспроизводства (Волобуев и др., 2001; Volobuev et al., 2002). Работы выполнялись в Петропавловско-Командорской, Тихоокеанской, Северо-Охотоморской рыбопромысловых подзонах.
Кета ежегодно встречалась во всех рыбопромысловых подзонах на протяжении всего периода наблюдений. Последостижения кетой, нагуливающейся в северо-западной части Тихого океана, генетически обусловленных морфофизиологических кондиций начинается ее преднерестовая миграция в направлении Охотского моря, куда она проходит в основном через Четвертый Курильский пролив (Бирман, 1985), а также через другие проливы, находящиеся между Четвертым и Фриза (Шунтов, 1994). Проводимые исследования позволили проследить зарождение миграционных потоков кеты в Петропавловско-Командорской подзоне, их развитие в Тихоокеанской подзоне (Северо-Курильской зоне) и завершение в Северо-Охотоморской подзоне (табл. 19).
Как следует из табл. 19, кета составляла основу уловов в Северо-Охотоморской подзоне и выступала в роли субдоминанта по отношению к другим видам лососей в Петропавловско-Командорской и Тихоокеанской подзонах. Ее наибольшие уловы на одно рыбопромысловое усилие в Петропавловско-Командорской подзоне наблюдались в июне, а в Тихоокеанской и Северо-Охотоморской
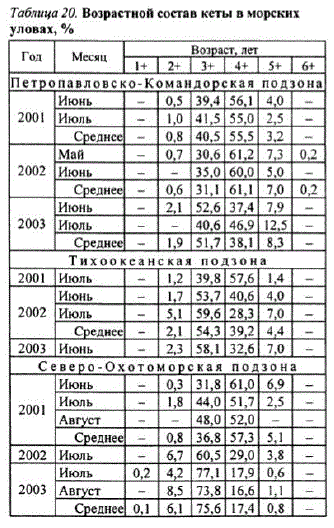
подзонах, как правило, в T5 2O. B. июле, что обусловлено L:NE= аолица 20 возрастной составкеты в морских
мой ее миграционного пути в уловах * периодморских преднерестовых миграций (см. табл. 19),
В морских уловах кета ными группами: от 1 + - до 5+ лет встречались ежегодно. Доминировали 4-летние (3+) и 5-летние (4+) особи, составлявшие в сумме от 87,5 до 100% уловов. Доля рыб в возрасте 21 и 5 лет не превышала 12,5%. Двух и 7-летние (6+) особи встречались единично, и их доля не превышала 0,2% За время исследований размеры кеты варьировали ственно, 59,5 см и 2,81 кг. В уловах практически всегда преобладали самки или соотношение полов было примерно одинаковым, доминирование самцов в уловах было от
Таблица 21. Биологические показатели кеты
Основные биологические показатели кеты подвержены как межгодовой. так и географической изменчивости. Например, кета с наименьшими средними значениями длины и массы тела была отмечена в 2002 г., а с наибольшими — в 2001 г., тогда как ежегодно наименьшими линейными и весовыми показателями характеризовалась кета, отлавливаемая в Северо-Охотоморской подзоне, наибольшими, как правило, кета Тихоокеанской подзоны. На протяжении исследований в течение одного сезона, как обычно, во всех подзонах в течение преданадромной миграции наблюдалось увеличение линейно-весовых размеров кеты, свидетельствующее о замещении в миграционном потоке ранней формы ее поздней темпоральной группировкой (см. табл. 21).

В Петропавловско-Командорской подзоне и Северо-Курильской зоне Тихоокеанской подзоны в уловах преобладала кета с гонадами на II—III и III стадиях зрелости. В Северо-Охотоморской подзоне отмечено преобладание кеты с гонадами на III—IV и IV стадиях зрелости (табл. 22). Это хорошо заметно по наиболее представительным данным 2001 г. В то же время в 2003 г. в уловах встречалось более 40% неполовозрелой кеты. Неполовозрелая кета среди мигрирующих к местам воспроизводства лососей в Охотском море встречалась и в прежние годы. В 1993 г. доля таких рыб составила 29%, а в 1994 г. — 38% (Волобуев и др., 1995). Среди неполовозрелых рыб (стадия зрелости гонад II) в эти годы преобладали 4-летки (3+): соответственно 62,8 и 63,5%. Большая часть (77,8—68,2%) неполово
Таблица 22. Соотношение кеты с гонадами на разных стадиях зрелости, %

зрелой кеты была представлена To || Meca Стадия зрелости самцами, причем основная ее масса встречалась в начале хода (июнь). В июле доля незрелых рыб понижалась до 8—10%. По результатам траловых съемок в зимний период неполовозрелой кеты в Охотском море Cpeне обнаружено (Радченко и др., численность очень низка (0,3%) (Шунтов, 1989). Наличие агрегаций :
неполовозрелых особей кеты в Охотском море в июне-июле, возMOKHO, обусловлено привлечением их шлейфом видоспецифичных феромонов и аттрактантов, которые выделяются массой идущих нанерест зрелых рыб этого вида. Значительные скопления неполовозрелой кеты (до 80%) в океане в районесеверных Курильских островов и в Северо-Охотоморская подзона OxoTcKoM море отмечались ранее
рый считал, что сезонные миграции (200 играют важную роль в реализации
В желудках кеты отмечены — каляниды, гиперииды, эвфаузииды, Июль гаммариды, рыбы (преимущественно светящиеся анчоусы), крылоногие моллюски, молодь кальмара, гребневики и щетинкочелюстные. Основу питания кеты практически всегда составляли ракообразные.
Как показано в табл. 23, спектр питания кеты был подвержен межгодовой и пространственной изменчивости. Например, в Петропавловско-Командорской подзоне в 2001 г. в желудках кеты наиболее часто встречались эвфаузиевые раки и гребневики, в 2002 г. — каляниды, а в 2003 г. — гиперииды и молодь рыб. Однако если в Петропавловско-Командорской подзоне в 2001 г. основу питания кеты составляли каляниды, то в Тихоокеанской и Северо-Охотоморской подзонах — гиперииды.
Таблица 23. Спектр питания кеты в северо-западной части Тихого океана и в Охотском море
Доля в пише, %
У большинства рыб степень наполнения желудков варьировала от 1 до 3 баллов. Доля особей с полным растянутым желудком (4 балла) обычно не превышала 10% и лишь в отдельных случаях достигала почти 20%. Наряду с этим во всех районах исследования наблюдались особи спустыми желудками и их доля варьировала от 11,8 до 67,3% (табл. 24). Столь широкий диапазон варьирования доли рыб, не имевших в желудках кормовых объектов, по-видимому, связан не сплохими условиями обитания, а с качественным составом пищи кеты, что ранее отмечали Н.В. Кловач с соавторами (1996). В нашем случае наибольшая доля рыб спустыми желудками наблюдалась в июле 2001 г. (67,3%), когда основу питания составляли гребневики и эвфаузииды, а наименьшая доля
(11,8%) — в 2002 г., когда основными компонентами в питании кеты были рыбы и калянусы (см. табл. 23, 24). Кроме того, степень наполненности желудков пищей зависит от времени, проведенного рыбами в сетях.
Таблица 24. Наполнение желудков кеты, %
Во всех исследованных районах проведены массовые анализы для оценки анатомических отклонений в развитии гонадкеты.
Все основные выявленные фенодевианты можно отнести каномалиям, могущим снижать репродуктивный потенциал: перехлесты, нахлесты, перекручивания семявыносящего протока, перетяжки, фрагментарное деление надоли, недоразвитость одной из гонади др. (Микодина, Пукова, 2002) (рис. 19—24).
Все указанные изменения морфологии гонад характерны в основном для самцов, встречаемость самок с подобными нарушениями развития половых желез невелика (Volobuev et al., 2002) (табл. 25). Для Петропавловско-Командорской подзоны доля самцов кеты саномалиями гонад составила 52,3%, самок — 0,6%. Основная доля самцов саномалиями развития гонаду кеты приходится на массовые возрастные группы — 3+ и 4+. В Северо-Курильской зоне доля самцов кеты с фенодевиантами семенников была также высокой — 67,6%, У самок фенодевианты яичников не отмечены.
Таблица 25. Доля особей кеты с аномалиями развития гонад, %

Рис. 19. Нормальная морфология половых желез тихоокеанских лососей: ст— самок, б — самцов
Рис.20. Перехлесты и перекруты в головной и хвостоной частях семенников кеты
Рис.2). «Морщинистая» поверхность семенников кеты, обусловленная множественными мелкими шрамами от небольших поверхностных очагов резорбцин генеративнойткани
Рис. 23. Левый семенник в виде яйцевидного образования с долей на поверхности, в правом-нахлести подвернутголовной конец
Рис.24. Яичники кеты с жировыми опухолями и гематомами
В Северо-Охотоморской подзоне доля рыб с аномальным развитием гонад оставалась высокой: у самцов она составила 65,8%. В Южно-Курильской зоне доля самцов с нарушениями развития семенников была еще выше, чем в более северных районах Тихого океана и Охотском море, — 84,8%.
Единичные аномалии в развитии гонадукеты отмечались ранее (Shershneva, Vvedenskaja, 2001). В большем количестве такие аномалии характерны для восточносахалинской кеты (Gгitsenko et al., 2001). Следует отметить, что по мере соматического ростагонад число фенодевиантов гонадрезко снижается — более чем на порядок ковремени захода лососей в реки. Возможно, наличие массовых отклонений в развитии семенников является следствием алломстрии роста половых желез, которые, по-видимому, в основном нивелируются в процессе трофического ростагонад.
Для проведения исследований по оценке эффективности работы ЛРЗ и идентификации JOCOce OTHOCITeJILHO районов их происхождения широко применяются термические методы массового мечения тихоокеанских лососей. Внедрение в середине 1990-х гг. на лососевых рыбоводных заводах Магаданской области прогрессивных методов массового термического маркирования североохотоморской кеты посредством нанесения меток на отолиты позволило в настоящее время приступить к исследованиям по определению районов морского нагула и распределения охотоморских и других дальневосточных стад.
Кета, имевшая на отолитах метки, нанесенные на ЛРЗ, в сборах встречалась ежегодно. Места ес поимок в северо-западной части Тихого океана и в Охотском море в 2001—2005 гг. приведены на рис. 25.
Основываясь на количестве рыб, меченых на ЛРЗ и затем пойманных в море, по формуле, предложенной Петерсоном и модифицированной Чепменом (Рикер, 1979), можно оценить численность кеты того или иного региона происхождения в море:
(M+1)(C+1)
R+1
где M — количество меченых рыб, экз.; С— объем пробы, взятой при исследо
ваниях, экз.; R — количество меченых рыб в пробе, экз.
Например, в результате расчетов численность рекрутов кеты североохотоморского происхождения в нагульных скоплениях в Петропавловско
Командорской и Тихоокеанской подзонах, которые должны были подойти на нерест в 2001 г., была оценена в 2724,7 тыс. рыб. Подход кеты в реки северного побережья Охотского моря в 2001 г. составил 1704,0 тыс. рыб, т.е. потери в период нагула и преднерестовой миграции в море оценены в 1020,7 тыс.- экз. (около 37%), и, по нашему мнению, в первую очередь, они связаны с общей смертностью лососей в море. Это может быть следствием болезней, выедания хищниками, промысла и т. д.
Рис. 25. Карта-схема мест поимок североохотоморской кеты, помеченной термическими метками: 62.05.2 — Петропавловско-Командорская подзона, 62.03.1 — СевероКурильская зона (тихоокеанская сторона); 65.05.1 — Северо-Охотоморская подзона; 65.05.4 Камчатско-Курильская подзона
Как показали наблюдения, проведенные в Притауйском районе Охотского моря (56—57° с.ш., 146—153° в.д.) на судах-дрифтероловах в 1993—1995 гг. (Волобуев и др., 1995), миграция кеты к побережью не ограничивается только прибрежными водами, а охватывает всю акваторию моря. Миграционные потоки кеты после преодоления ею Курильских проливов распределяются веерообразно. В Притауйском районе интенсивный ход ранней кеты в реки отмечен уже в первых числах июня, когда уловы на стандартную пелагическую сеть длиной 50 м достигали 10—12 экз. при температуре поверхности воды 1,6—2,5°C. Средняя масса рыб в течение июня составила 2,5—2,7 кг. Во II декаде июля она заметно увеличилась, и в уловах встречались особи массой 2,7—3,2 кг, что свидетельствовало о начале преднерестовой миграции поздней кеты.