Актуальные проблемы акклиматизации горбуши в Белом море
Тихоокеанский лосось – горбуша широко расселенный вид в бассейне Тихого океана, заходящий для нереста в реки как азиатского, так и американского побережий. Среди дальневосточных лососей рода Oncorhynchus, представленного 6-ю хорошо различающимися видами, горбуша по общим представлениям исследователей, является наиболее специализированным видом, обладающим по сравнению с другими видами рядом уникальных особенностей: этот вид имеет среди тихоокеанских лососевых самый короткий 2-х летний жизненный цикл; у него отсутствуют пресноводные формы; после выхода из бугров молодь задерживается в пресной воде наиболее короткое время (1-2 месяца); снижен «инстинкт дома» (хоминг); кариологическая эволюция по типу робертсоновских транслокаций завершена (все хромосомы двуплечие, 2n=52, NF=104); и, наконец, вид слабо дифференцирован на внутривидовом уровне (несмотря на широкое распространение, у него отсутствуют подвиды), в то же время вид имеет сложную популяционную структуру (генетически различающиеся летние и осенние подходы, локальные стада, генерации четных и нечетных лет).
Уникальным среди лососевых является и ранее появление зародышевых клеток и быстрое развитие гонад, которые первоначально у всех особей формируются как яичники. Передифференцировка пола завершается перед выходом личинок из гнезд (Персов, 1975). Среди этих особенностей особое внимание привлекают репродуктивно изолированные поколения горбуши смежных лет, т.е. заходящие на нерест в четные и нечетные годы. Особенности «четной» и «нечетной» линий весьма существенны и заключаются в следующем: в четные годы производители в среднем подходят на нерест позднее; четная линия, как правило, малочисленнее, число хромосом в четной линии стабильно и равно 2n=52, NF=104; все хромосомы двуплечие; в нечетный оно варьирует 2n=54, 53, 52 (причем последнее наиболее редко), при NF=104, т.е. в кариотипе не все хромосомы двуплечие, отличаются эти линии и по степени генетической межпопуляционной дифференциации (в четной линии она выше). Таким образом, степень экологических и генетических различий между этими линиями формально достигает видовых, хотя существенных морфологических особенностей (на которых обычно основываются таксономические построения) у этих линий нет. Скорее всего, они могут рассматриваться как виды-двойники.
Впервые о возможности акклиматизации дальневосточных лососей на европейском Севере еще в XIX веке говорил академик Карл Бэр. Попытки акклиматизации горбуши (а также кеты) в разных регионах мира предпринимались неоднократно. Целенаправленное и масштабное вселение горбуши в бассейн Белого моря началось с 1956 года, продолжалось до 1980 г. Необходимость его была связана с бедностью ихтиоценоза в этом регионе, а также падением численности самого ценного вида – семги.
Всего было перевезено свыше 200 млн. икринок, которые затем доинкубировались на рыбоводных заводах. Икра завозилась с Южного Сахалина. Вселенцы широко расселились и оказались способными к естественному нересту и нагулу. Самовоспроизводящиеся популяции горбуши были отмечены даже в Норвегии (Berg, 1977). На Белом море горбуша была введена в промышленный лов (Рис. 1). Начиная с 1960 г., на рыбозаводах проводилась инкубация икры, собранной от местных производителей. Однако возвраты производителей были нестабильны и постепенно стали ничтожными (уловы колебались от 144 тыс. экз. в 1973 г. до единичных в 1983 г.). При этом выяснилось, что четные и нечетные линии ведут себя по-разному: уловы в нечетные годы значительно превышали таковые в четные. На протяжении этого периода акклиматизации проводились интенсивные научные исследования (Агапов, 1986; Азбелев и др., 1961; Азбелев, Яковенко, 1963; Бакштанский, 1963; Bakshtansky, 1980; Бирман, 1985; Дягилев, Маркевич, 1979; Камышная, Смирнов, 1981; Карпевич и др., 1991; Сурков, Суркова, 1977; Яковенко, 1992 и др.). Было установлено, что неудачная акклиматизация связана, в основном, с несоответствием температурных условий в южной части нативного ареала горбуши, откуда завозилась икра, и суровых условий европейского Севера (Дягилев, Маркевич, 1979).
В 1985 г. начался второй этап акклиматизации вселенца: была завезена небольшая (около 2,5 млн.) партия икры нечетной линии с севера ареала из Магаданской области, а в 1998 г. четной линии в таких же количествах. В результате численность горбуши в нечетные годы достигает несколько десятков тысяч экз. (34-156 тыс.), хотя и подвержена значительным колебаниям. Необходимо подчеркнуть, что значительные колебания численности характерны для горбуши не только Белого моря, но и для обоих смежных поколений в нативном ареале. Нужно отметить также, что с 1985 г. рыбоводные заводы инкубируют икру от местных производителей лишь эпизодически. Завоз икры четной линии в 1998 г. увеличения уловов не дал. Учитывая, что со времени последнего завоза этой лини прошло уже 3 поколения, 2004 г. был решающим в отношении эффективности проведенного вселения этой линии. Прогнозы на этот год полностью подтвердились, заход рыб на нерест в большинстве рек (Кереть, Умба) был единичным, что неудивительно, принимая во внимание ее отличительные особенности, о которых было уже сказано. Падение уловов и постепенное исчезновение горбуши после первого этапа вселения и сложные экономические условия в нашей стране привели к падению интереса исследователей к изучению последствий вселения. Научные изыскания возобновились лишь в последние годы после второго этапа акклиматизации. Основное внимание направлено на проведение мониторинга за состоянием популяции. В соответствии с разработанной программой исследования проводятся силами сотрудников ЗИН РАН, СПбГУ, Институтом биологии КНЦ РАН, ПИНРО, СевПИНРО при поддержке Минпромнауки, а также сотрудниками Института общей генетики РАН.
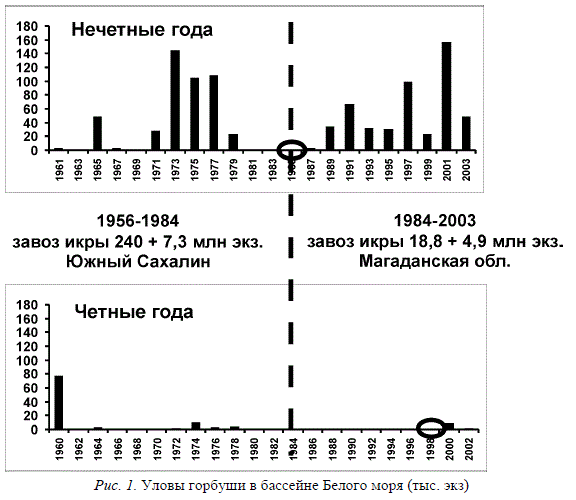
Рис. 1. Уловы горбуши в бассейне Белого моря (тыс. экз)
Исследования проводились в рамках проблемы «Структурно-функциональная оценка экосистем Белого моря, как основа развития многоплановой марикультуры и повышения ее биопродуктивности» (головное учреждение ЗИН РАН). Далее будут рассмотрены результаты работ по изменению структуры популяции горбуши, изменению ее морфологического облика и основам генетических преобразований в формирующейся популяции.
В 2001-2003 гг. исследования были проведены сотрудниками разных институтов в разных регионах бассейна Белого моря. Специалисты ЗИНа и СПбГУ работали в районе о. Средний (губа Чупа) и анализировали динамику нерестового хода производителей, их морфологическую характеристику, питание и развитие половых желез. Один из выводов этих работ состоит в том, что в новом регионе обитания горбуша сохранила присущие ей в нативном ареале основные характеристики нерестового хода, такие как волнообразная динамика миграции, увеличение к концу хода доли самок и более крупных производителей и другие. Установлено (Рис. 2), что к началу августа в море горбуша практически полностью перестает питаться, относительная масса пищеварительного тракта с пищей снизилась до 0,5%. В то же время относительная масса яичников последовательно увеличивалась и достигла к началу августа ¼ общей массы тела, что характерно для самок горбуши непосредственно перед нерестом. Дефинитивных размеров достигала и масса ооцитов. Таким образом, все исследованные производители потенциально готовы были нереститься до 15 сентября, т.е. в сроки, которые согласно проведенным ранее исследованиям (Дягилев, Маркевич, 1979), являются благоприятными для последующего выживания и развития молоди. Это дает основания полагать, что в популяции беломорской горбуши на протяжении 9 поколений нечетной линии в результате отбора сохранились в основном ранненерестящиеся особи. Нужно отметить, что именно такой путь был предложен рядом ученых при селекции производителей на рыбоводных заводах для создания ранненерестующего стада (Карпевич и др., 1991).
Нечетная линия беломорской горбуши демонстрирует адаптивные сдвиги и по ряду других морфобиологических признаков (линейные размеры, масса тела, сроки массовой нерестовой миграции, сроки пребывания эмбрионов и личинок в буграх, плодовитости самок). Обнаружено, что в новом ареале нерестовая миграция взрослых рыб в реки начинается несколько раньше. На протяжении нерестового хода преобладают самки. У самок беломорской горбуши заметно увеличивается плодовитость. Вследствие того, что в новом ареале нерест происходит раньше, чем в нативном ареале, а скат молоди из рек из-за позднего прогрева воды весной, наоборот, происходит позже, молодь дольше остается в нерестовых буграх. Мальки горбуши уже в районе нерестилищ и во время ската начинают питаться, что им не свойственно в реках Магаданской области.
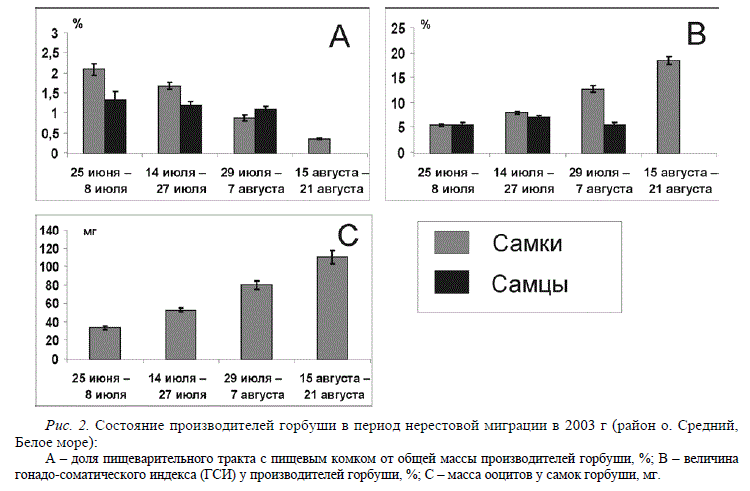
Рис. 2. Состояние производителей горбуши в период нерестовой миграции в 2003 г (район о. Средний, Белое море): А – доля пищеварительного тракта с пищевым комком от общей массы производителей горбуши, %; В – величина гонадо-соматического индекса (ГСИ) у производителей горбуши, %; С – масса ооцитов у самок горбуши, мг.
Сравнительные исследования пластических признаков беломорской популяции четной и нечетной линий демонстрируют сдвиги в ее морфологическом облике. Еще на первом этапе акклиматизации было отмечено в нечетной линии (Суркова, 1970), что уже через 4-5 поколений беломорская горбуша стала иметь меньше позвонков и чешуй в боковой линии, у нее уменьшилась голова и соответственно ее пропорции. Результаты дискриминантного анализа 17 пластических признаков четной и нечетной популяций (Гордеева, 2003), проведенного на последнем этапе акклиматизации, свидетельствуют о существенных сдвигах в их морфологическом облике, причем сдвиги более выражены у четной линии и, по-видимому, инадаптивны, в то время как нечетная линия демонстрирует процесс адаптации к новым условиям.
Остеологические, более консервативные, признаки горбуши в нативном ареале характеризуются сравнительно малой изменчивостью, однако в наших более ранних исследованиях (Горшков, Дорофеева и др., 1979; Горшков, 1979) была отмечена слабая тенденция к клинальной изменчивости некоторых признаков, связанных с костями этмоидального отдела черепа и имеющих таксономическое значение.
Сравнительные исследования остеологических признаков беломорской горбуши нечетной линии позволяют судить об изменениях в форме некоторых костей. При этом, как и в некоторых внешнеморфологических признаках (Суркова, 1970), формирующаяся популяция беломорской горбуши проявляет сходство не только с донорской популяцией, но и несет черты, свойственные другим, часто более южным, популяциям. Так, форма супраэтмоида беломорской горбуши имеет черты, с одной стороны, сходные с горбушей из реки Большой на Камчатке (очертания наружных боковых краев кости), а с другой, с горбушей из реки Анадырь (малая глубина каудальной вырезки). В то же время сошник у рыб из реки Кереть сходен по форме с сошником рыб из Приморья, а язычная кость по основным параметрам (форма головки и рукоятки) сходна по форме с язычной костью рыб из рек Камчатки (Рис. 3).
Необходимо отметить также, что перечисленные особенности беломорской горбуши относятся лишь к модальной группе рыб. В целом же все остеологические особенности отличаются большой изменчивостью, и у рыб из реки Кереть можно найти почти весь спектр остеологических признаков, свойственных рыбам, обитающим в различных районах естественного ареала. Есть основания предполагать, что изменения всех морфологических признаков наступают сравнительно быстро, т.к. Суркова (1970) отмечала их уже в 4-5-м поколениях.
В серии исследований, посвященных генетической структуре беломорской горбуши (Гордеева, 2003; Гордеева и др., 2003), выявлены изменения по сравнению с донорской популяцией (редукция числа гетерозигот и аллельного разнообразия, а также разнообразия гаплотипов мтДНК). В целом уровень генетических изменений существенно выше в нечетной линии, чем в четной, что на фоне успешного естественного воспроизводства у первой, может свидетельствовать о процессах адаптации. В то же время на основании имеющихся данных можно предполагать, что неутешительные итоги акклиматизации четной линии горбуши в Белом море могут быть связаны с ее более узкой специализацией по сравнению с нечетной линией (хотя с уверенностью об этом следует судить лишь по прошествии еще 5-6 генераций II-го этапа вселения четной линии). Об этом же свидетельствуют данные акклиматизации горбуши в бассейне Атлантического океана на американском континенте. Попытки акклиматизации нечетной линии на некоторое время давали положительные результаты, хотя полная натурализация произошла только при образовании жилой формы в Великих озерах после интродукции нечетной линии горбуши из тихоокеанского бассейна в 1955 г. Возврата от четной линии вселения обычно не было (Hoard, 1991).
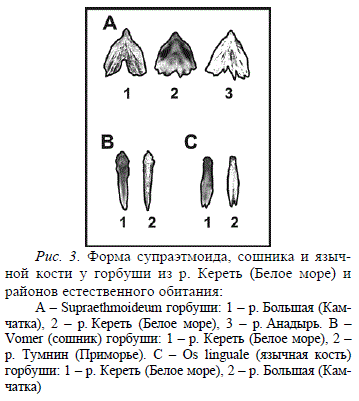
Рис. 3. Форма супраэтмоида, сошника и язычной кости у горбуши из р. Кереть (Белое море) и районов естественного обитания: А – Supraethmoideum горбуши: 1 – р. Большая (Камчатка), 2 – р. Кереть (Белое море), 3 – р. Анадырь. В – Vomer (сошник) горбуши: 1 – р. Кереть (Белое море), 2 – р. Тумнин (Приморье). С – Os linguale (язычная кость) горбуши: 1 – р. Кереть (Белое море), 2 – р. Большая (Камчатка)
Наибольший интерес с теоретической и практической точек зрения представляет вопрос о завершении натурализации горбуши в Белом море, т.е. возникновения устойчивой самовоспроизводящейся популяции. Этот вопрос можно решить только при продолжении всестороннего мониторинга. Сравнительно стабильные уловы после последнего этапа интродукции нечетной линии и намечающиеся генетические и морфобиологические адаптации дают основания говорить о возможности процесса натурализации. В ходе первого неудачного периода акклиматизации при практически непрерывном завозе икры и существенной подпитке популяции рыбозаводами падение уловов произошло примерно через 13 поколений. На втором этапе прошло уже 9 поколений (при крайне малом завозе икры и без подпитки рыбозаводами инкубацией икры от местных производителей). Тем не менее, падения уловов в нечетные годы не наблюдается. Относительный успех 2-го этапа может быть обусловлен 3-мя неравноценными факторами. На первом месте, по сей видимости, стоит удачный выбор донора, затем возможность нарастания численности местного стада (в том числе происходящего и от первого периода акклиматизации). И, наконец, возможно играет существенную роль наступление периода потепления.
Выводы
«Нечетная» линия горбуши, акклиматизированная в Белом море, на протяжении 9 поколений сохраняет устойчивые подходы производителей, что может свидетельствовать о формировании ранненерестующей популяции горбуши. «Нечетная» линия горбуши в Белом море обнаруживает существенные морфологические и генетические сдвиги, которые на фоне устойчивых уловов позволяют судить о протекающих процессах адаптации в формирующейся беломорской популяции горбуши.
О завершении процесса натурализации и о месте горбуши в экосистеме Белого моря можно будет судить только после всестороннего и регулярного анализа данных, полученных в результате мониторинга нескольких популяций беломорской горбуши на протяжении не менее 15 поколений.
Е.А. ДОРОФЕЕВА1, А.П. АЛЕКСЕЕВ2, В.Г. КУЛАЧКОВА1, О.В. ЗЕЛЕННИКОВ3, Т.С. ИВАНОВА3
1 Зоологический институт РАН, Санкт-Петербург
2 Межведомственная ихтиологическая комиссия, Москва
3 Санкт-Петербургский государственный университет, Санкт-Петербург