ПИЩЕВЫЕ ОТНОШЕНИЯ МОЛОДИ АТЛАНТИЧЕСКОГО ЛОСОСЯ С ТУВОДНЫМИ ВИДАМИ РЫБ
Наиболее сходны спектры питания молоди лосося и хариуса (табл. 99),
особенно у молоди лосося с сигом и бычком-подкаменщиком. При оценке
сходства питания молоди лосося и хариуса на видовом уровне (на примере
ручейников) СП-коэффициенты понижаются, но остаются самыми
высокими по сравнению с другими видами рыб (Мартынов, 1983). На
сходство питания хариуса и молоди лосося обращали внимание многие
исследователи (Никольский и др., 1947; Владимирская, 1957; Заболоцкий,
1959 и др.). Наибольшее сходство в составе пищи наблюдается между
сеголетками лосося и хариуса (Сидоров, 19796; Мартынов, 1983), хотя
оно остается высоким и у старших рыб (1+... 3+) при более широком видовом
составе кормовых объектов в пище молоди лосося (Сидоров, Шубина,
1990).
Спектры питания молоди лосося и бычка-подкаменщика перекрываются
за счет крупных личинок двукрылых и поденок, молоди лосося и
сига - личинок двукрылых, ручейников и веснянок. При сопоставлении с
другими видами рыб на уровне отрядов и семейств потребляемых объектов
перекрывание значительно меньше (СП-коэффициенты не более 20).
Высокая степень перекрывания спектров питания видов не может
служить показателем напряженности пищевых отношений между ними,
а, скорее, свидетельствует о потенциальных возможностях ее возникновения
при низком уровне развития кормовой базы, высоких плотностях
рыб или полном совпадении микростаций обитания.
Сеголетки лосося на ранних стадиях находятся в межгалечниковых
пространствах нерестового грунта, а на более поздних занимают полосу
прибрежий с преобладающими глубинами 20—40 см и повышенными скоростями
течения, придерживаясь каменистого грунта. Старшевозрастные
пестрятки перемещаются на центральные участки порогов. В системах
Таблица 99. Степень сходства пищи рыб р. Щугор (СП-коэффициенты; по: Мартынов,
1983)
ТаЫе99. Similarity degree of food offish in the Shugor River (after: Martynov, 1983)
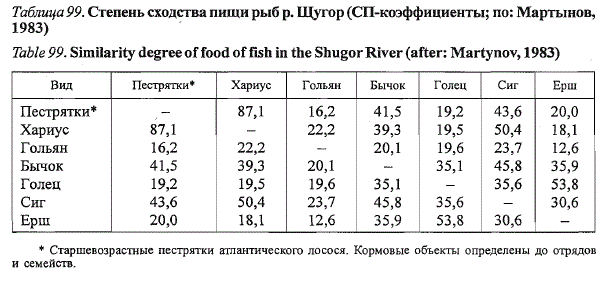
* Старшевозрастные пестрятки атлантического лосося. Кормовые объекты определены до отрядов
и семейств.
крупных лососевых рек отмечаются сезонные миграции старшевозраст-
ных пестряток в мелкие притоки основной реки (Веселов, Калюжин, 2001;
устное сообщение О.В. Букова для р. Илыч и СВ. Красикова - для р. Мезень).
Перед ледоставом молодь возвращается в основную реку. В малых
лососевых реках молодь более территориальна и, по-видимому, до ската
в море не совершает сколько-нибудь протяженных миграций. Пестрятки
зимуют на порожистых участках рек под камнями (Владимирская, 1957;
Шустов, 1983; Веселов, Калюжин, 2004).Молодь атлантического лосося
зимует под камнями на порожистых участках рек на Североамериканском
континенте (Cunjak, 1988).
Пространственная структура хариуса более лабильна. Сеголетки придерживаются
прибрежной полосы, но в отличие от лосося предпочитают
участки с более низкими скоростями течения, обычно на границе зарослей
околоводной растительности {Nordosmia), где мальки стайками
держатся в толще воды. В результате стации обитания сеголеток лосося и
хариуса, спектры питания которых в основном перекрываются, расходятся
практически полностью.
Старшевозрастная молодь хариуса перемещается на более глубокие
участки порогов, где перекрываются ее стации обитания и старшевозрас-
тных пестряток лосося, имеющих сходные размеры. Но даже при совместном
питании на одном и том же участке их микростации существенно
различаются (рис. 85). Молодь лосося занимает охотничью позицию на
каком-нибудь камне и, совершая кормовые броски от дна до поверхности,
в основном облавливает дно и придонные горизонты воды. После каждого
броска пестрятка возвращается в исходную позицию на камне. Хариус
держится в толще воды и, постоянно перемещаясь над охотничьей
позицией пестрятки, облавливает верхние слои воды. Картину совместного
питания старшевозрастной пестрятки (судя по размерам в 10 -12 см
в возрасте 3+) и одноразмерного с ней хариуса мы наблюдали в первой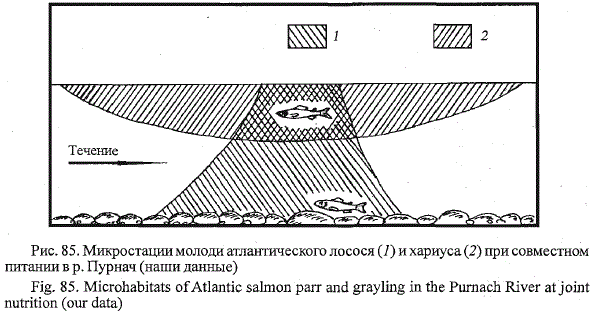
Рис. 85. Микростации молоди атлантического лосося (/) и хариуса (2) при совместном
питании в р. Пурнач (наши данные)
Fig. 85. Microhabitats of Atlantic salmon parr and grayling in the Purnach River at joint
nutrition (our data)
половине августа 1983 г. в р. Пурнач, притоке Поноя. Интересно отметить,
что пестрятка не проявляла по отношению к хариусу агрессии, почти
сталкиваясь с ним в зоне перекрывания охотничьих участков. Сопоставив
рис. 85 и 86, можно оценить особенности гидрологических характеристик
на охотничьих участках пестрятки и молоди хариуса (поверхностная
скорость течения на кормовой стации сравниваемых видов составляла
0,6-0,7 м/с). Если в облавливаемой пестряткой зоне скорость течения
варьировала практически от нулевых значений в охотничьей точке
примерно до 0,6 м/с у поверхности, то хариус, облавливавший верхний
горизонт воды, постоянно находился в потоке со скоростями течения
примерно 0,4-0,6 м/с. Скорости потока, близкие к нулевым значениям,
отмечены в охотничьих точках пестряток лосося в р. Порья (Бакштанс-
кий, Нестеров, 1983). Рассмотренный нами отдельный случай совместного
питания одноразмерных пестрятки и молоди хариуса в ламинарном
потоке наглядно демонстрирует расхождение их микростаций обитания.
В естественных условиях гидротопографические ниши подвержены значительной
изменчивости (Веселов, Калюжин, 2001), и конкретные формы
взаимодействия видов в различающихся условиях также могут варьи-
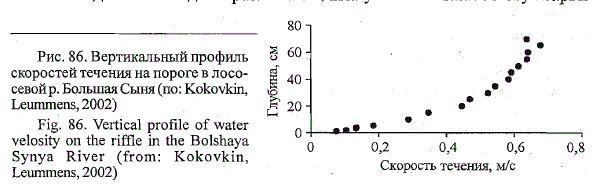
Рис. 86. Вертикальный профиль
скоростей течения на пороге в лососевой
р. Большая Сыня (по; Kokovkin,
Leummens, 2002)
Fig. 86. Vertical profile of water
velosity on the riffle in the Bolshaya
Synya River (from: Kokovkin,
Leummens, 2002)
235
ровать. Но большая привязанность пестряток лосося к грунту, а молоди
хариуса—к толще воды, скорее всего, будет сохраняться во всех случаях.
О питании молоди хариуса преимущественно в толще воды можно судить
по высокой доле в ее рационе куколок и имаго ручейников (Шубина,
1986). Значительный видовой спектр ручейников, потребляемых пест-
рятками (Сидоров, Шубина, 1990), определяется адаптацией последних к
питанию в более широком, по сравнению с хариусом, диапазоне параметров
гидротопографической ниши.
Помимо хариуса, совместно с пестрятками лосося на порожистых участках
различных лососевых рек обитают гольян, бычок-подкаменщик,
голец усатый, кумжа, елец. Как было сказано выше, у гольяна можно выделить
две экологические группы: потамофильную, обитающую в прибрежье,
и реофильную, использующую медиаль перекатов. В последней
группе выделяются называемый нами одиночный «пестряточный» гольян,
поведение которого даже в деталях копирует поведение сеголеток лосося,
и реофильный стайный.
Потамофильная форма гольяна исключается из конкурентных отношений
с молодью лосося как по составу питания (доминируют водоросли),
так и по занимаемой стации. Реофильный стайный гольян питается на
порогах и активно перемещается по ним, используя придонный слой воды
(Веселов, Калюжин, 2001). Наиболее близок по занимаемой микростации
и характеру поведения к младшевозрастным пестряткам лосося «пестряточный
» гольян. Подразделение гольяна на три группы (потамофиль-
ный стайный, реофильный стайный и реофильный одиночный) отчетливее
всего прослеживается в р. Варзуге, где отсутствуют такие облигат-
ные литореофилы, как бычок-подкаменщик и голец усатый. По-видимому,
реофильные формы гольяна, в какой-то мере занимают их экологические
ниши и могут ограничивать нишу пестряток. В той или иной мере
экологическая дифференциация гольяна наблюдалась нами и в других
лососевых реках.
Бычок-подкаменщик и голец усатый обитают на порогах совместно с
пестрятками лосося. Их микростации ограничиваются придонным слоем
воды, не превышающим высоту тела. Совершая перемещения, эти виды
не теряют тактильного контакта с грунтом, а при опасности укрываются
под камнями. В наибольшей мере их стации совпадают со стациями пес-
тряток-сеголеток. Гольцы, бычки, «пестряточные» гольяны и сеголетки
лосося могут укрываться под соседними камнями. О топографической
совместимости стаций этих четырех видов свидетельствует их одновременное
присутствие в уловах электроловильным аппаратом (Martynov et
al., 1994). При увеличении размеров пестряток и расширении облавливаемой
ими топогидравлической ниши перекрывание микро стаций с об-
лигатными литореофилами уменьшается. Аналогичными пищевым поведением
и стациями характеризуется голец усатый. Остроту пищевых
отношений между гольцом усатым и бычком-подкаменщиком в значи-
236
тельной степени нивелирует расхождение спектров их питания, которое
определяется различиями в строении ротового аппарата.
Стации сига и ерша приурочены к элементам русел лососевых рек с
замедленными скоростями течений. В связи с этим перекрывание спектров
питания этих двух видов с молодью лосося не приводит к пищевой
конкуренции.
Кумжа в системах лососевых рек заселяет мелкие притоки, которые не
используются молодью лосося. В р. Йоканьге речная форма кумжи занимает
нишу хариуса, и отношения ее молоди и лосося во многом должны
быть сходны с взаимоотношениями молоди лосося и хариуса.
Известно, что наибольшая пищевая конкуренция может возникать
между видами рыб, представляющих разные фаунистические комплексы
и питающихся сходными группами организмов (Никольский и др., 1947;
Никольский, 1965). На НВУ атлантического лосося в реках Севера России
наименьшие СП-коэффициенты наблюдаются у видов одного фаунисти-
ческого комплекса. У хариуса, гольяна, бычка-подкаменщика и гольца усатого
(предгорный бореальный фаунистический комплекс) пищевые спектры
перекрываются за счет второстепенных объектов питания, в то время
как основу пищи составляют беспозвоночные разных систематических
групп. Наглядным примером коадаптации видов одного фаунисти-
ческого комплекса со сходной экологией могут служить подкаменщик и
голец усатый, которые ведут придонный образ жизни и держатся совместно.
Однако морфобиологические особенности этих рыб приводят к тому,
что основу пищи бычка составляют крупные объекты, в частности Diptera
n/d, а гольца — мелкие личинки хирономид. У видов со сходной экологией
- выходцев из разных фаунистических комплексов — спектры питания
значительно перекрываются за счет основных объектов питания. Близка
степень сходства пищи ерша и гольца усатого, сига и хариуса, сига и бычка.
Однако пищевые отношения между вышеуказанными парами видов
не реализуются в виде конкурентных в связи с различиями их макростаций:
ерш и сиг обитают на участках рек с замедленным течением, а голец
усатый, хариус и бычок - на порогах.
Большим сходством отличаются спектры питания молоди лосося и
хариуса. При совместном обитании на порожистых участках рек данные
виды, принадлежащие к разным фаунистическим комплексам, избегают
пищевой конкуренции за счет расхождения микростаций обитания.
Смотрите также