ЭКОЛОГИЯ МОЛОДИ АТЛАНТИЧЕСКОГО ЛОСОСЯ В РЕКАХ СЕВЕРА РОССИИ
Пестрятки. В связи с отсутствием достоверных половых различий в
длине и массе одновозрастной ювенильной молоди лосося (р > 0,05) данные
по неполовозрелым самцам и самкам, приводимые далее в таблицах,
объединены. Увеличение массы гонад созревающих самцов-пестря-
ток на Севере России наблюдается в первой половине августа. В этот
период по коэффициентам зрелости самцов можно подразделить на три
группы. Гонады ювенильных самцов (группа I) находятся на второй стадии
зрелости, и их доля от массы тела не превышает 0,1 %. Гонады самцов
II группы визуально относятся ко второй стадии зрелости, но их го-
надо-соматический индекс превышает 0,1 % и соответствует таковому
самок. Самцы этой группы, видимо, из-за слабого развития гонад не в
состоянии созреть к сентябрю и принять участие в нересте, в связи с чем
они также отнесены нами к ювенильным. И, наконец, Ш группа самцов
имеет гонады на третьей и четвертой, включая промежуточные, стадиях
развития. Их гонадо-соматический индекс превышает 1,0 % и в середине
августа у отдельных особей может достигать более 10,0%; Самцы этой
группы рассматриваются нами как карликовые.
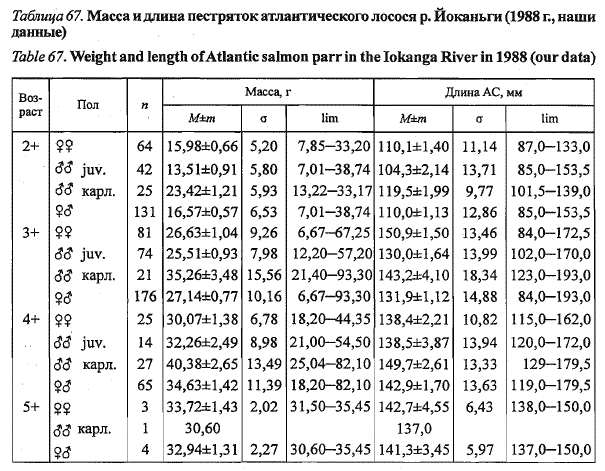
В р. Иоканьге часть самцов-пестряток созревает по карликовому типу
на третьем году жизни (табл. 67). Молодь характеризуется высокой изменчивостью
размерно-весовых показателей. При достоверных различиях
средних значений длины и массы (р<,0,05) крайние показатели особей в
смежных возрастных группах в значительной мере перекрываются. В уловах
на крючковую снасть доминировали рыбы в возрасте 3+ и 2+ (46,8 и
34,8 % соответственно). Пестрятки младшей возрастной группы (2+) были
представлены практически равным количеством самцов и самок. Среди
рыб старших возрастных групп (3+ и 4+) дрля самцов возрастала до 54 и
Таблица 67. Масса и длина пестряток атлантического лосося р. Йоканьги (1988 г., наши
данные)
Table 67. Weight and length of Atlantic salmon parr in the Iokanga River in 1988 (our data)
 Возраст
Возраст63 % соответственно. В старшей возрастной группе (5+) доминировали
самки, что может быть обусловлено случайными причинами и связано с
небольшим объемом исследовательской выборки.
Коэффициенты зрелости большинства самок-пестряток йоканьгского
лосося не выходили за пределы 1,0 %. Исключение составила одна самка
в возрасте 2+, длиной по Смитту 114,5 мм и массой 16,6 г, не включенная
в табл. 67. Ее гонады были увеличены в размерах и находились на III
стадии зрелости, гонадо-соматический индекс составил 1,69 % и оказался
сопоставимым с таковым карликовых самцов. Не исключено, что данная
самка могла созревать по карликовому типу.
В р. Пурнач, правобережном лососевом притоке р. Поной, в уловах
мальковым неводом и удочкой пестрятки были представлены пятью возрастными
группами. Доминировали рыбы в возрасте 2+, что объясняется
их высокой долей в уловах мальковым неводом, который был малоэффективным
при облове участков с быстрым течением и крупновалунным
грунтом, где держались старшевозрастные пестрятки. Молодь в возрасте
4+ встречалась единично (табл. 68). Судя по возрастному составу уловов
пестряток, смолтификация молоди лосося в р. Пурнач и скат в море в
основном происходят в возрасте трех и четырех лет.
Таблица 68. Коэффициенты зрелости и упитанности пестряток атлантического лосося
р. Йоканьги (наши данные), %
Table 68. Gonad-somatic and body condition coefficients of Atlantic salmon parr in the Iokanga
River (our data), %
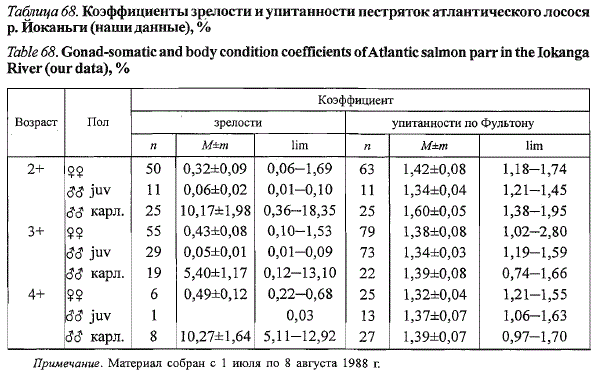
Примечание. Материал собран с 1 июля по 8 августа 1988 г.
В р. Варзуге достоверных половых различий в длине и массе тела неполовозрелых
пестряток не выявлено (табл. 69). Линейные размеры и
масса карликовых самцов практически во всех возрастных группах превышали
соответствующие показатели ювенильных рыб. В возрастных
группах пестряток 0+ и 1+ преобладали самки, а среди старших рыб -
самцы (доля самцов в 1982-1985 гг. в возрастных группах 0+ - 3+ составила
47,0; 44,4; 52,2 и 58,3 % соответственно). Повышение доли самцов
среди старших рыб связано с созреванием части самцов по карликовому
типу с последующей задержкой в реке. Созревание карликовых самцов в
р. Варзуге отмечено на втором году жизни рыб (в возрасте 1+). В этом
возрасте, по нашим данным, в среднем за ряд лет созревает 4,2 % самцов.
Среди старшевозрастных пестряток доля карликов увеличивается. В возрастной
группе 2+ в среднем созревает 29,3 % самцов, а в группе 3+ -
47,6 %. Коэффициенты зрелости карликовых самцов увеличиваются с
приближением нерестового периода. За все годы исследований средний
показатель коэффициента их зрелости в конце августа составил 11,3 %
при колебаниях от 3,5 до 16,2 %.
В р. Чаваньге пестрятки лосося представлены шестью возрастными
группами (табл. 70). Часть самцов начинает созревать по карликовому
типу на третьем году жизни. В возрастных группах от 4+ и выше встречены
только карликовые самцы. Высокая изменчивость биологических показателей
рыб во многом связана с тем, что в таблицу включены материалы,
собранные на протяжении длительного периода времени. Так, коэффициент
зрелости карликового самца в возрасте 5+, выловленного
Таблица 69. Средние показатели длины и массы пестряток атлантического лосося
р. Варзуги (по: Драганов и др., 1990)
Table 69. Length and weight of Atlantic salmon parr in the Varzuga River (after: Draganov
at al., 1990)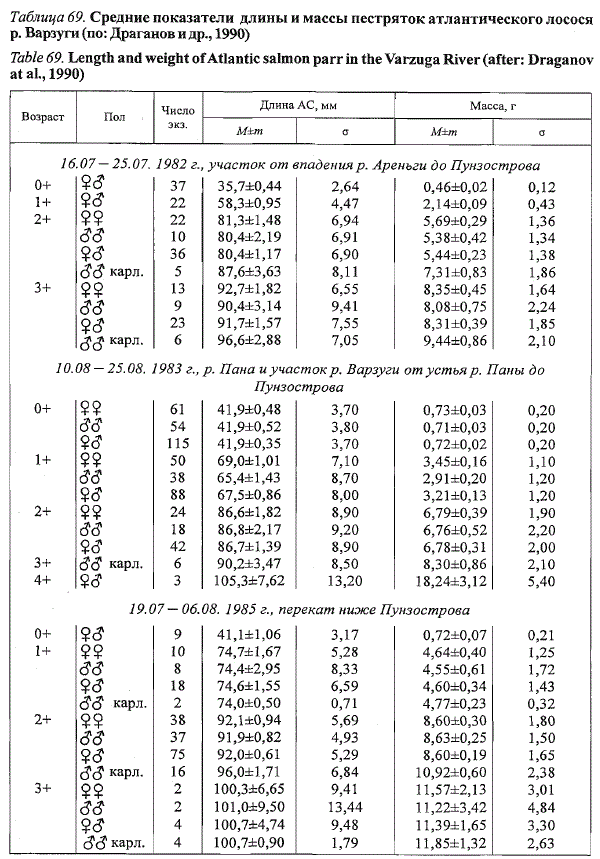
Таблица 70. Коэффициенты зрелости, масса и длина пестряток атлантического лосося
р. Чаваньги (наши данные)
Table 70. Gonado-somatic coefficients, weight and length of Atlantic salmon parr in the
Chavanga River (our data)
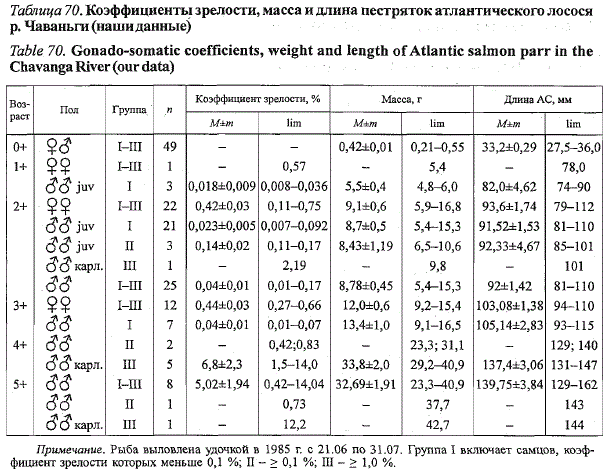
Примечание. Рыба выловлена удочкой в 1985 г. о 21.06 по 31.07. Группа I включает самцов, коэффициент
зрелости которых меньше 0,1 %; II'- > 0,1 %; Ш - > 1,0 %.
29 июня, составил 0,73 %, а соответствующий показатель одновозраст-
ного самца, выловленного 25 июля, - 12,2 %.
Одновозрастная молодь лосося из различных уральских притоков Печоры,
отловленная в близкие сроки, характеризуется сходными показателями
длины и массы тела (табл. 71). Молодь из р. Пижмы, тиманского
притока Печоры, значительно превосходит по размерам одновозрастных
рыб из правобережных притоков, причем различия увеличиваются с возрастом,
и трехлетки (2+) более чем в два раза по массе превосходят своих
ровесников, выловленных в близкие календарные сроки в уральских притоках
Печоры.
В отличие от лососевых рек Кольского полуострова в притоках Печоры
длина и масса карликовых самцов меньше, чем у ювенильных ровесников,
что наглядно демонстрируют материалы по пестряткам рек Под-
черем и Печорская Пижма (табл. 72, 73).
Состав молоди лосося на нерестилищах в бассейне Печоры до ската в
море характеризуется равным соотношением полов (табл. 74). После ската
в море среди оставшихся на нерестилищах старшевозрастных пестряток
начинают преобладать самцы, которые созревают по карликовому
типу. Созревание самцов в возрасте 1+ отмечено для верховий Печоры,
Таблица 71. Средние показатели массы и длины пестряток атлантического лосося в
бассейне Печоры (по: Мартынов, 1983; Martynov et al., 1994 с дополнениями)
Table 71. Average weight and length of Atlantic salmon parr in the Pechora River basin (after:
Martynov, 1983; Martynov et al., 1994 with additions)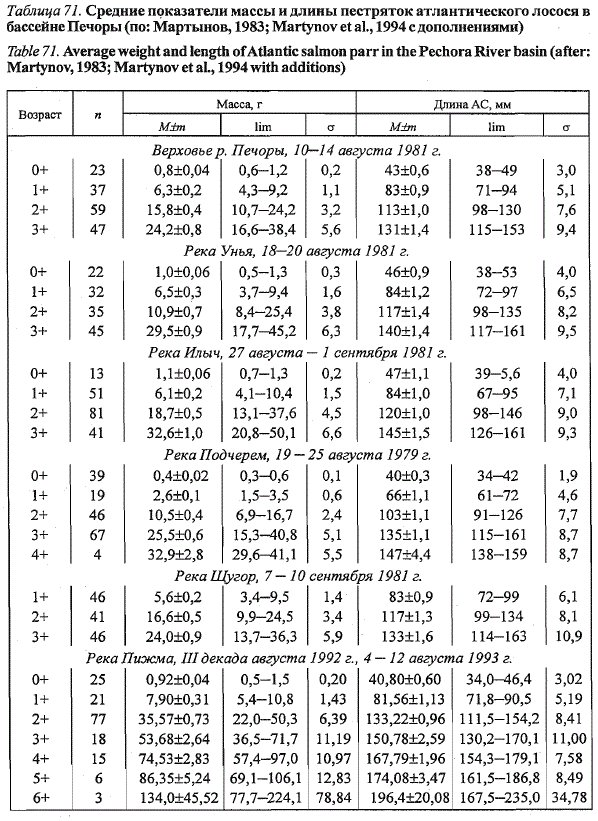
Таблица 72. Коэффициенты зрелости, масса и длина пестряток атлантического лосося р. Подчерем (наши данные)
Table 72.Gonado-somatic coefficients, weight and length of Atlantic salmon parr in the Podcherem River (our data)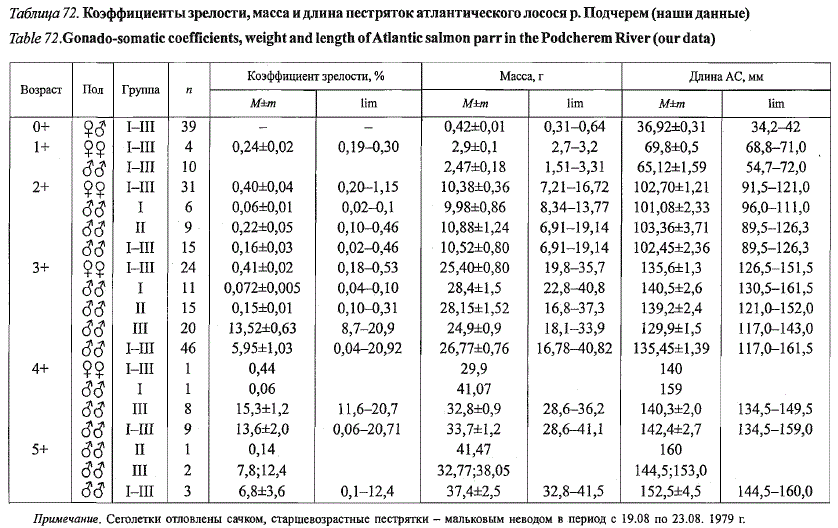
Примечание. Сеголетки отловлены сачком, старшевозраетные пестрятки - мальковым неводом в период с 19.08 по 23.08. 1979 г.
Таблица 73. Коэффициенты зрелости, масса и длина пестряток атлантического лосося р. Печорской Пижмы (1992 -1993 гг. (наши
данные)
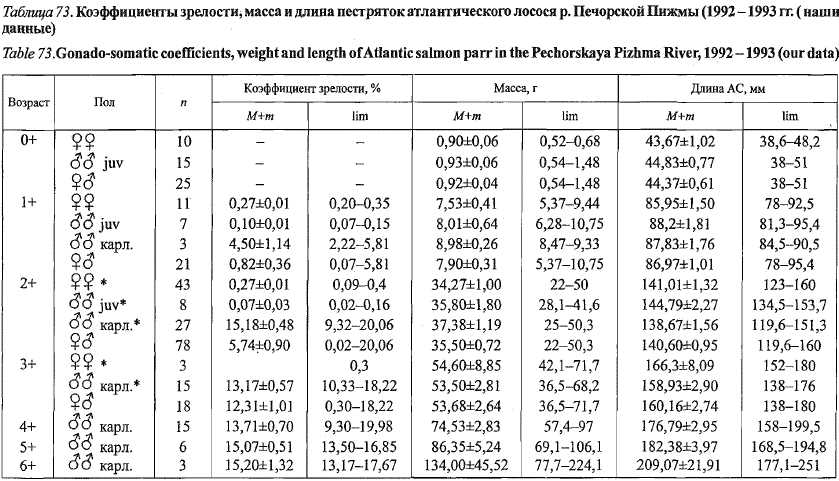
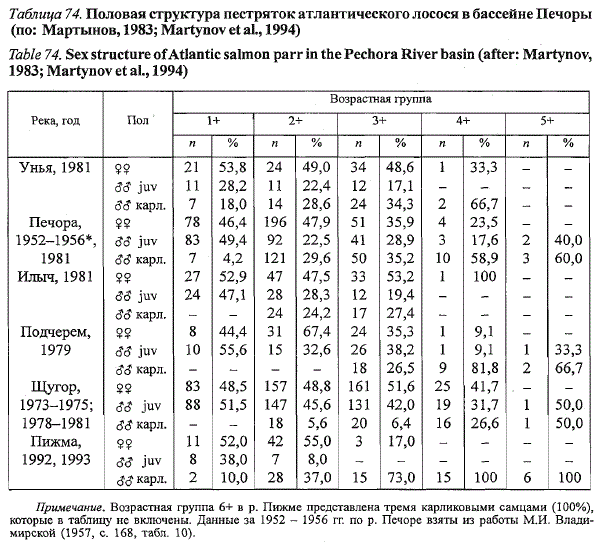

Table 75.Gonado-somatic coefficients, weight and length of Atlantic salmon parr in the Pechorskaya Pizhma River, 1992 -1993 (our data)
* Коэффициенты зрелости в возрастных группах 2+ определены у 38 самок; 5 ювенильных самцов и 25 карликовых самцов; 3+ - у одной самки и 14
карликовых самцов; «-» - данные отсутствуют.
Таблица 74. Половая структура пестряток атлантического лосося в бассейне Печоры
(по: Мартынов, 1983; Martynov et aL, 1994)
Table 74. Sex structure of Atlantic salmon parr in the Pechora River basin (after: Martynov,
1983; Martynov et aL, 1994)
Примечание. Возрастная группа 6+ в р. Пижме представлена тремя карликовыми самцами (100%),
которые в таблицу не включены. Данные за 1952 - 1956 гг. по р. Печоре взяты из работы М.И. Владимирской
(1957, с. 168, табл. 10).
рек Унья и Пижма, где более интенсивен прогрев воды в летний период.
Созревание карликовых самцов в лососевых реках Приполярного Урала
начинается в возрасте 2+. Наибольшая часть ювенильных самцов созревает
в год, предшествующий скату молоди в море: в уральских притоках -
в возрасте 3+, в таманской Пижме - 2+. Среди пестряток, возраст которых
превышает доминантный возраст покатников, преобладают карликовые
самцы. Их доля в р. Пижме в возрастных группах 4+ и выше достигает
100 %. Наибольший возраст отмечен нами у карликового самца из
р. Кожим (бассейн Усы). Возраст отнерестившегося самца, имевшего признаки
лошания, напоминавшие таковые у крупных производителей, датируется
как 7+ (рис. 70). Очевидно, что карликовые самцы, возраст которых
превышает максимальный возраст покатников, принимали участие
в размножении неоднократно. С другой стороны, значительная часть
карликовых самцов младшего возраста, принимавших участие в нересте,
весной следующего года скатывается в море (см. ниже).
Рис. 70. Чешуя карликового самца атлантического
лосося р. Кожим. Выловлен 22 октября
19 81 г. в нижнем течении реки. Масса тела 23 5 г,
длина по Смиту 278 мм, возраст 7+ лет
Fig. 70. Scale of precocious male of Atlantic
salmon caught in the Kozhim River 22 October
1981. Weight 235 g, fork length 278 mm, age 7+
Покатники. Речной период жизни
молоди атлантического лосося завершается
катадромной миграцией и сменой
гипоосмотической среды обитания
на гиперосмотическую. Переход молоди
из рек в море связан со сложными
морфо физиологическими изменениями
в организме рыб, рассматриваемыми
в специальной литературе (см. Вар-
навский,1990). Нас в первую очередь
интересуют межпопуляционные особенности биологической структуры
покатников и ее адаптивная значимость.
Покатная миграция молоди лосося в реках Восточного Мурмана начинается
в первой половине июля, когда вода прогревается до 8,2-11,5 °С,
а заканчивается в конце июля — первой половине августа при температуре
воды 13,5-21,0 °С (табл. 75). Массовый скат приходится на вторую-
третью декады июля и длится 7 - 1 0 дней при прогреве воды до максимальных
значений. В р. Сидоровке средняя длительность ската молоди
составила 23+6 дней при межгодовых колебаниях от 12 до 33 дней (и = 3
года). В р. Варзина в 1978 г. молодь скатывалась в течение 35 дней при
миграции основного контингента рыб с 22 по 30 июля (Кузьмин, 1985).
При многовозрастной структуре (шесть групп - 2+... 7+) средний возраст
покатников в малых реках Восточного Мурмана колеблется от 3,4 до
4,2 года. Средняя длина покатников р. Сидоровки изменялась от 15,2 до
15,7 см, масса- от 35,3 до 42,6 г (Кузьмин, 1985). Обращает на себя внимание
сходство структуры речного периода жизни лосося, определенной
по чешуе рекрутов, с возрастной структурой покатников.
Скат молоди лосося в р. Пече, притоке р. Туломы, в 1983 г. начался в
первой декаде июня, а закончился в конце первой пятидневки июля. Два
покатника были отловлены 17 июня. При контрольном облове 3 июля,
когда температура воды в реке составила 18 °С, покатной молоди не обнаружили.
В уловах удочкой присутствовали только пестрятки в возрасте
1+ и 2+. Таким образом, основной скат молоди лосося в р. Пече в 1983 г.
проходил в течение месяца и завершился в первой пятидневке июля.
Скатившись из нерестовых рек в Нижнетуломское водохранилище, покатники
лосося попадают в среду, существенно отличающуюся по темпе-
Таблица 75. Продолжительность ската молоди атлантического лосося и температура
воды в реках Сидоровка (1974 -1976 гг.) и Варзина (1978 г.) (по: Кузьмин, 1985)
Table 75. Duration of downstream migration of Atlantic salmon smolts and water temperature
in the Sidorovka (1974 -1976) and the Varzina (1978) Rivers (after: Kuzmin, 1985)
ратурным условиям от реки. Однако покатная молодь довольно быстро преодолевает
водохранилище: в 1983 г. подход покатников к плотине Нижне-
туломской ГЭС наблюдался 12 июня при температуре воды 5 °С, а в 1985 г.
первые покатники были отловлены удочкой у плотины ГЭС 27 июня при
температуре воды 8 °С. Следует отметить, что 12 июня 1983 г. температура
воды в нижнем течении р. Печи составила 11 °С. По данным В.В. Азбеле-
ва (1960), скат молоди из притоков Туломы в 1959 г. начался при температуре
воды 6 °С и был наиболее интенсивным при 12,5 - 16,5 °С.
Покатная молодь туломского лосося концентрировалась у плотины
перед турбинами Нижнетуломской ГЭС в поверхностном слое воды в
зоне ее кругового движения у правого берега (рис. 71). Вместе с током
воды стайки молоди многократно совершали циклические перемещения,
то приближаясь вплотную к плотине, то удаляясь от нее на 40 - 50 м.
В 1983 г. наибольшую численность, судя по количеству
всплесков и частоте поклевок, стайки
покатной молоди имели в период с 16 по
30 июня, после чего плотность скоплений снизилась,
хотя серебристая молодь ловилась на
удочку и спиннинг до 25 июля. В небольшом
количестве покатная молодь лосося встречается
перед плотиной Нижнетуломской ГЭС в
ее верхнем бьефе в течение всего лета и осе
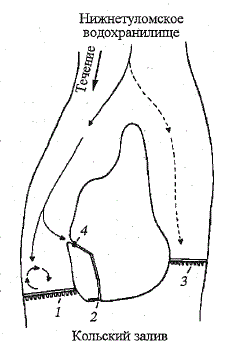
-
Рис. 71. Схема миграции покатной молоди атлантического
лосося в районе гидросооружений Нижнетулом-
скойГЭС.
1 - плотина ГЭС; 2 - рыбоход; 3 - водослив; 4 - рыбоучет-
ная ловушка. Стрелками показано направление движения рыб
Fig. 71. The scheme of downstream migration of Atlantic
salmon smolts near the dam of the Lower Tuloma
hydroelectric power station.
1 - dam; 2 - fish way; 3 - water sink; 4 - fish registering trap.
Arrows indicate fish migration direction
нью. Так, 18 октября 1983 г. в рыбоходе после его закрытия было выловлено
4 экз., а 18 октября 1985 г. - 5 экз. крупной молоди с хорошо выраженными
признаками посеребрения. По устному сообщению ихтиолога
управления «Мурманрыбвод» Т.Ф. Мишуковой, осенью в отдельные годы
в рыбоходе обнаруживали до 300 экз. серебристой молоди лосося.
Большинство покатников благополучно преодолевают гидросооружения
Нижнетуломской ГЭС в отличие от Верхнетуломской. По данным
Д.С. Павлова с соавт. (1999а), «...Смолты лосося без повреждений проходят
Нижнетуломскую ГЭС, а на Верхнетуломской от действия перепада
давления их гибнет около 15 %», а у оставшейся в живых молоди «.. .реакция
на различного рода раздражители... начинала проявляться только
по истечении 30 мин после прохождения турбины» (с. 139, 151).
Средняя масса покатной молоди лосося (без учета рыб, выловленных
поздней осенью в водохранилище и рыбоходе) в 1983 г. составила
33,1±0,78 г при средней длине 153,4±1,13 мм (и = 147), в 1985 г. - 30,6±
±0,83 г при средней длине 144,1±1,19 мм (п = 128). Покатники имели
хорошо выраженную серебристую окраску тела, хотя у многих из них просматривались
поперечные темные полосы и красные пятна, свойственные
окраске пестряток. Основная масса рыб скатывалась в море в возрасте
3+ и 4+ лет при среднем возрасте 3,9 года (табл. 76).
Покатная молодь, задерживающаяся до осени перед плотиной Нижнетуломской
ГЭС и в рыбоходе, отличается крупными размерами, обусловленными
большим летним приростом (Мартынов, Куценко, 1985). Отдельные
особи покатников могут оставаться на зимовку в водохранилище.
Число склеритов на чешуе покатников, отловленных нами 10 июля
1983 г. перед плотиной ГЭС, колебалось от 0 до 8 (в среднем 4,1), а у рыб,
задержавшихся в рыбоходе до 15 октября, - от 13 до 19 (в среднем 15,3).
Выживаемость задержавшейся перед гидросооружениями молоди, по-
видимому, невысока. Из 356 производителей туломского лосося, исследованного
нами в 1985 г., лишь у 12 прирост в речной зоне роста чешуи был
представлен более чем восемью склеритами, а их максимальное число в
приросте, равное 11, отмечено у одного. Судя по числу склеритов в приросте
речной зоны, рыбы, задержавшиеся перед гидросооружениями до
осени, среди возвращающихся на нерест производителей отсутствовали.
В р. Иоканьга покатная миграция молоди обычно происходит в июле
и продолжается в течение 30-35 дней. Пик миграции приходится на вторую
декаду июля. Подход первых экземпляров покатной молоди к устью
реки отмечен при температурах воды 11,8-13,9 °С. Массовый выход покатников
в море обычно происходит при прогреве воды в реке до 15,0-
17,8 °С. В теплом 1989 г. массовый подход покатников к устью реки наблюдался
при прогреве воды до 24,8 °С и проходил на две недели раньше
обычных сроков (Зубченко и др., 1991; Долотов, 2006).
Средние показатели массы и длины тела покатников йоканьгского лосося
незначительно различаются по годам (табл. 77). Например, показате-
Таблица 76. Масса и длина покатников атлантического лосося р. Туломы, выловленных
удочкой (наши данные)
Table 76. Weight and length of Atlantic salmon smolts in the Tuloma River (our data)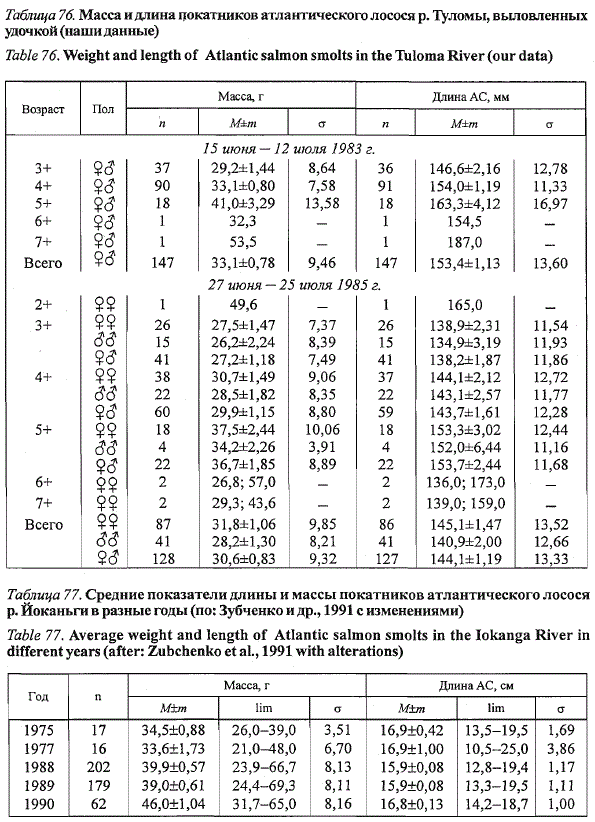
Таблица 77. Средние показатели длины и массы покатников атлантического лосося
р. Йоканьги в разные годы (по: Зубченко и др., 1991 с изменениями)
Table 77. Average weight and length of Atlantic salmon smolts in the lokanga River in
different years (after: Zubchenko et al., 1991 with alterations)
Год
ли средней длины рыб изменяются в пределах сантиметра. Более значительные
различия массы тела могут быть связаны с уровнем накопления и
траты резервных питательных веществ в разные по условиям нагула годы.
Средняя за ряд лет наблюдений масса покатников составила 40,4 г при
средней длине тела 16,0 см (Зубченко и др., 1991). Возрастная изменчивость
средних размеров покатников несколько выше межгодовой, но в целом
невелика (табл. 78). Меньшие различия также характерны для показателей
средней длины разновозрастных покатников: в 1988 г. у рыб из наибольшей
(5+) и наименьшей (3+) возрастных групп различались на 0,7 см.
Половые различия в размерах одновозрастных покатников не выявлены.
Среди покатной молоди доминируют самки, доля которых в 1988 г.
составила 57,5 % (см. табл. 78), а в среднем за все годы наблюдений -
57,6 % (Зубченко и др., 1991), причем среди старшевозрастных рыб она
увеличивается. В 1988 г. среди рыб в возрасте 3+ самки составили 50,0 %;
4+ - 57,3 %; 5+ - 68,4 %, а в среднем за все годы наблюдений 55,0; 58,0 и
64,5 % соответственно (Зубченко и др., 1991). Возрастная изменчивость
полового состава покатников отрицательно коррелирует с таковой у пес-
тряток, что связано с созреванием части ювенильных самцов по карликовому
типу с последующей задержкой и повышенной смертностью в реке.
Среди покатников йоканьгского лосося отмечены рыбы в возрасте от
двух (2+) до шести (6-) лет (табл. 79). Рыбы крайних возрастных групп
представлены единичными экземплярами и могут отсутствовать в исследовательских
пробах в отдельные годы. Доминируют особи в возрасте
4+. В 1988 г. средний возраст покатников составил 4,03 года, за весь
период наблюдений - 3,88. Средняя продолжительность речного периода
жизни, определенная по чешуе покатников, несколько превышает соответствующий
показатель, определенный по чешуе производителей.
В р. Поной покатная миграция молоди лосося начинается в третьей
декаде июня, ее пик приходится на первую, а окончание - на третью декаду
июля (Неклюдов, 1990). По материалам М.Н. Неклюдова, любезно
предоставленным в наше распоряжение, за три года наблюдений (1984,
1986, 1987 гг.) скат молоди в среднем продолжался 33,7±0,7 дня. Средняя
масса покатников по годам изменялась от 22,54 до 26,49 г, а длина —
от 133,5 до 138,3 мм при среднегодовых значениях 23,97±1,26 г и
135,8±1,39 мм (табл. 80). Достоверные половые различия в размерно-
весовых показателях покатников не выявлены (р > 0,05), хотя как в пределах
возрастных групп, так и в целом прослеживается тенденция к доминированию
самцов. В смежных возрастных группах средние длина и масса
покатников различаются в пределах 1 см и 2 г, а в крайних возрастных
группах (3+ и 5+ за 1984 и 1987 гг.) разница составляет 1,7 см и 6,6 г.
Рыбы в возрасте 2+ и 6+ не учитывались ввиду их малочисленности.
В среднем за четыре года наблюдений среди покатников понойского
лосося преобладали самки (52,3±3,25 %) и доминировали рыбы в возрасте
3+ и 4+ лет. Средний возраст скатывающейся молоди за 1984-1987 гг.
182
Таблица 78. Средние показатели массы и длины покатников атлантического лосося
р. Иоканьги в разных возрастных группах (по: Зубченко и др., 1991 с дополнениями)
Table 78. Average weight and length of the Atlantic salmon smolts of different age groups in
the Yokanga River (after: Zubchenko et al., 1991 with additions)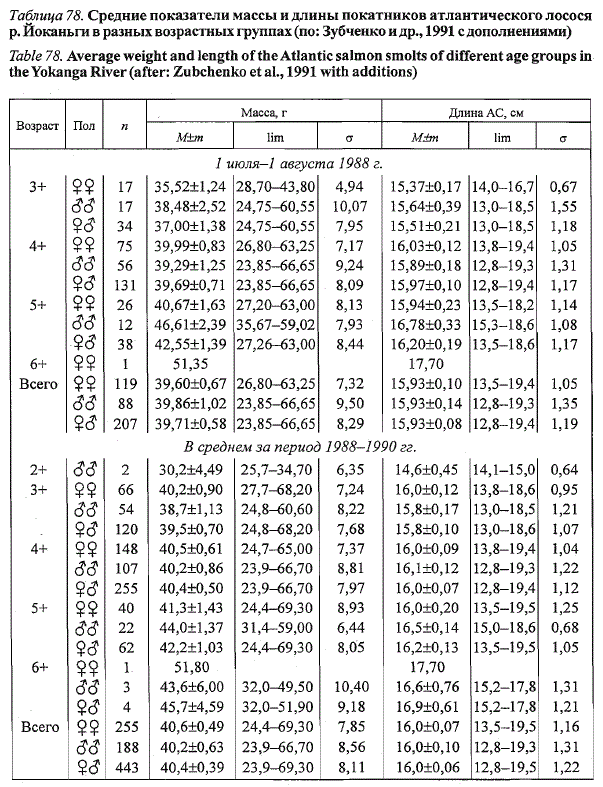
Примечание. Материал за 1988-1990 гг. собран и определен возраст рыб М.Н. Неклюдовым.
Таблица 79. Возрастной состав покатников атлантического лосося р. Йоканьги
(по: Зубченкоидр., 1991 с дополнениями; орудие лова-удочка), %
Table 79. Age structure of Atlantic salmon smolts in the Iokanga River, % (after: Zubchenko
et al., 1991 with additions)
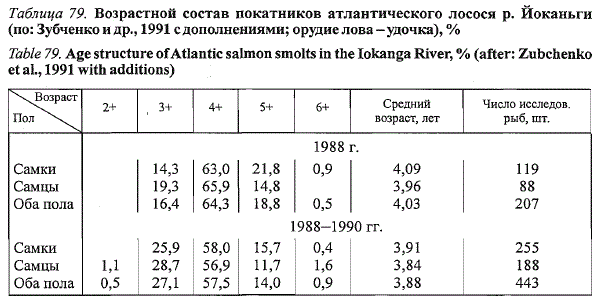
составил 3,6 года (2+ - 0,7; 3+ - 44,2; 4+ - 49,7; 5+ - 5,2; 6+ - 0,2 %;
расчет по табл. 80). Самцы жили в реке несколько дольше, чем самки ( в
среднем 3,64 и 3,57 года соответственно).
В течение миграции биологические показатели покатников закономерно
изменяются (Неклюдов, 1990). Скат старшевозрастных рыб приурочен
к первой половине покатного периода, доля младшевозрастных
рыб увеличивается к концу ската.
В р. Чаваньге в 1985 г. покатная миграция молоди лосося продолжалась
33 дня: с 16 июня по 18 июля. При средней длине 123,4 мм средняя
масса покатников составила 19,1 г (табл. 81) и почти в два раза превысила
соответствующие показатели покатников в соседней р. Варзуге (см.
ниже). Среди скатывавшейся в море молоди преобладали самки (61,4 %).
Среди самцов встречались особи с гонадами на III — VI стадиях зрелости,
что свидетельствует об их нересте предшествовавшей осенью по карликовому
типу. Доля таких рыб возрастала от 14,3 % среди самцов в возрасте
3+ до 25,0 % в возрасте 4+ и в среднем среди всех скатывавшихся в
море самцов составила 19,3 %. Показатели длины и массы ювенильной
молоди достоверно не отличались. Масса бывших карликовых самцов
была больше почти на 3 г, а длина на 1 см по сравнению с соответствующими
показателями ювенильной молоди, что говорит о более высоком
темпе роста карликовых самцов.
В р. Варзуге покатная миграция молоди лосося начинается по окончании
пика весеннего половодья и проходит в течение примерно одного
месяца при снижающемся уровне воды (Мельникова, 1959а, б, 1970; Ве-
селов, Казаков, 1998; Веселов, Калюжин, 2001). По данным М.И. Мельниковой,
покатная миграция варзугской молоди начинается при 9,6 °С,
Л.М. Нусенбаум (1953) - при достижении температуры воды 12 - 13 °С,
а пик ската- при 14,5 °С; Р.В. Казаков с соавт. (1992) наблюдали начало
Таблица 80. Масса и длина покатников атлантического лосося р. Поной (по: Неклюдов,
1990 с дополнениями)
Table 80. Weight and length of Atlantic salmon smoits in the Ponoy River (after: Nekljudov,
1990 with additions)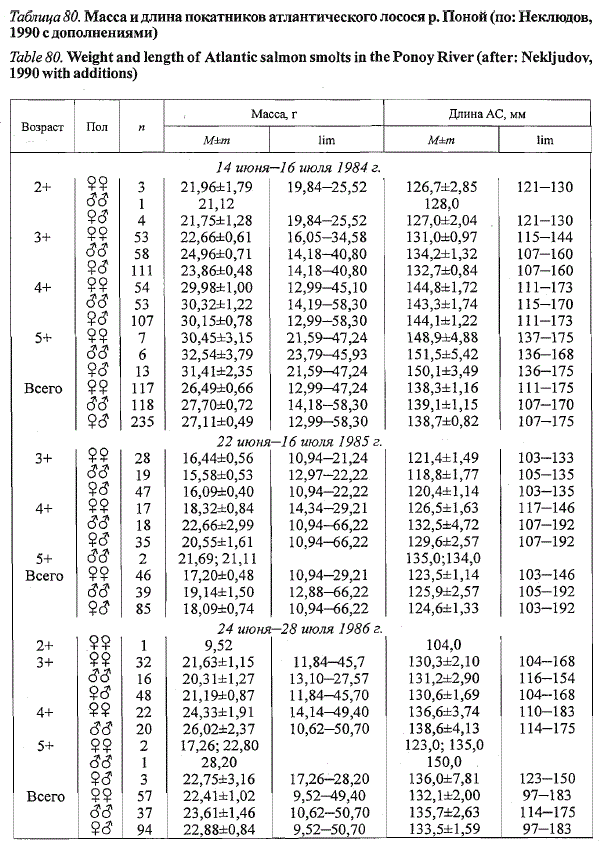
Таблица 81. Коэффициенты зрелости, масса и длина покатников атлантического лосося
р. Чаваньги (наши данные)
Table 81. Gonad-somatic coefficients, weight and length of the Atlantic salmon smolts in the
Chavan'ga River (our data)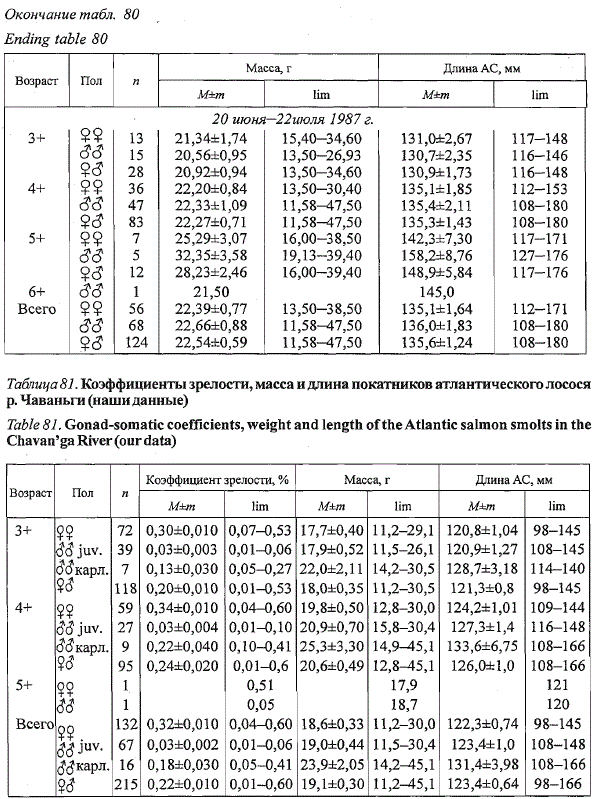
Примечание. Рыба отловлена удочкой в нижнем течении р. Чаваньги в период с 16.06 по 18.07
1985 г.
ската молоди 14 июня 1988 г. при температуре воды 11,5 °С, а в 1989 г. -
на полмесяца раньше при 13 °С. По их же данным, пик ската молоди в
1988 г. наблюдался 17-18 июня при температуре воды 15,5 °С, аего окончание
пришлось на конец июня (1988 г.) и первую декаду июля (1989 г.)
при прогреве воды до 17,3 °С. По нашим данным, покатная молодь лосося
встречалась в нижнем течении Варзуги (промысловый участок «Коло-
ниха») при температуре воды от 10,0 до 20,3 °С (1984 г.). Максимальные
скопления молоди перед стенкой рыбоучетного заграждения, свидетельствующие
о пике ската, зарегистрированы во второй половине июня при
температуре воды 15-17 °С. По нашим наблюдениям, скат молоди вар-
зугского лосося начинается в первой декаде июня и завершается к середине
июля, что совпадает с данными других авторов.
Достоверные половые различия по длине и массе у покатников вар-
зугского лосося, так же как и у пестряток, отсутствуют (табл. 82). Различия
же в средних показателях длины и массы рыб из смежных возрастных
групп достоверны (р < 0,05) и составляют 0,5-1,3 см и 2-3 г соответственно.
Среднегодовая разница между крайними возрастными классами (1969,
1984,1986,1988 гг.) достигает по длине 1,6 см, а по массе - 4,36 г.
В целом среди покатной молоди преобладают самки. Их среднегодовая
доля составила 51,2±2,27 % {п = 8 лет) при межгодовых колебаниях от
41,1 до 61,0 % (наши данные; Мельникова, 1959а, 1979; Казаков и др.,
1992). У покатников, как и у пестряток, доля самцов увеличивается в старших
возрастных группах: 2+ - 41,8 %, 3+ - 48,0 % и 4+ - 60,0 %. В сопоставимых
возрастных группах (1+ пестрятки, 2+ покатники и т. д.) более
низкая доля самцов среди покатников свидетельствует об их повышенной
смертности в реке в связи с созреванием по карликовому типу. Часть
варзугских карликовых самцов скатывается в море. По нашим данным,
они отличаются от ювенильных самцов внешним видом гонад (стадия
зрелости III—VI) и коэффициентом зрелости, равным таковому ювенильных
самок. В 1985 г. их доля составила 5,5 % от всей проанализированной
молоди, или 12,9 % от самцов.
Сопоставление наших данных с результатами других исследователей
(Мельникова, 1959, 1970; Казаков и др., 1992) показало, что средние годовые
величины длины и массы покатников слабо варьируют по годам.
За 11 лет исследований в среднем они составили 10,8±0,23 г при межгодовых
колебаниях от 10,2 до 12,7 г и 10,8±0,19 см при колебаниях от 9,9
до 12,3 см.
Половых различий в возрастном составе покатников нами не выявлено.
Средний возраст мигрирующей в море молоди мало меняется по годам
(табл. 83) и составляет 2,79±0,07 года при межгодовых колебаниях от
2,38 до 3,01 года (п = 8 лет).
После ската в море с конца июня до начала августа молодь лосося встречается
в прибрежных уловах беломорской сельди (Мельникова, 1959а; Аз-
белев, 1960). За это время на чешуе постсмолтов формируется в среднем
Таблица 82. Длина и масса покатников атлантического лосося р. Варзуги (по: Драганов
и др., 1990 с дополнениями)
Table 82. Weight and length of the Atlantic salmon smolts in the Varzuga River (after:
Draganovatal.,1990)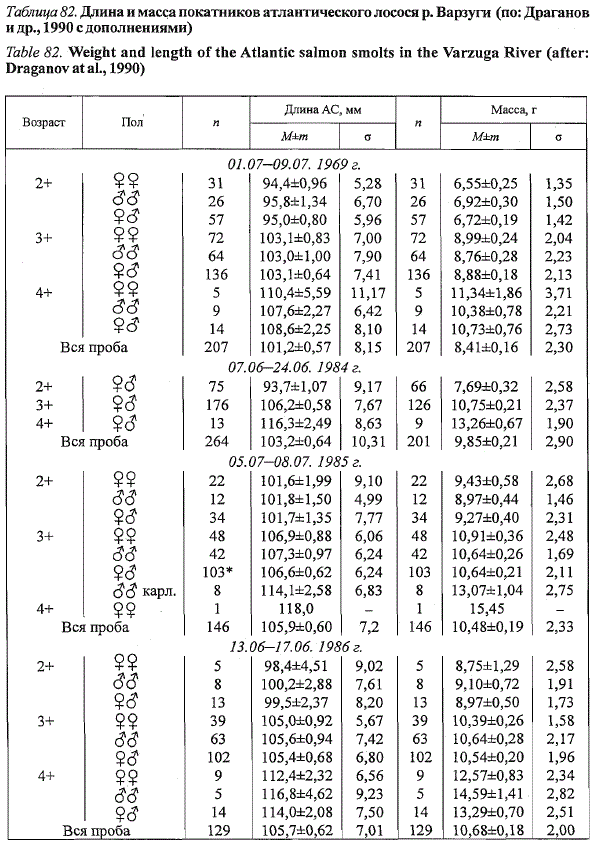
* Включены рыбы, пол которых не определялся.
Таблица 83. Возрастной состав покатников атлантического лосося р. Варзуги, %
Table 83. Age structure of the Atlantic salmon smolts in the Varzuga River, %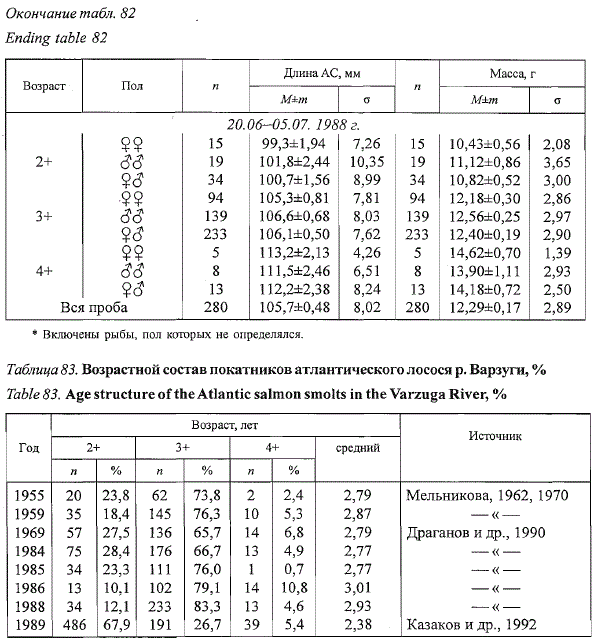
Казаков и др., 1992
12,5 склерита (колебания 6-18), которые по своей ширине занимают промежуточное
положение между узкими речными и широкими морскими
склеритами, а по числу соответствуют склеритам в переходной зоне на
чешуе производителей.
Данные о скате молоди печорского лосося немногочисленны (Владимирская,
1957; Vladimirskaya, 1959; Мартынов, 1983), что объясняется техническими
трудностями при сборе материалов в условиях многоводных
рек. Расхождение между средней продолжительностью parr-периода лосося
верховий Печоры, определенного по чешуе производителей (3,8 года),
и молоди на нерестилищах (доминанта 2+) М.И. Владимирская объясня-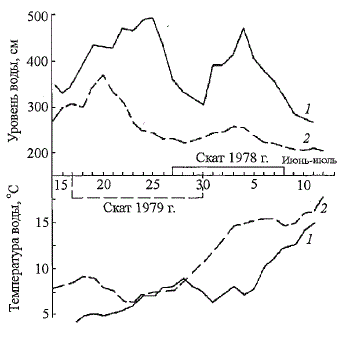
Рис. 72. Сроки и гидрологические
условия ската молоди атлантического
лосося р. Щугор в 1978 (7)
и 1979 (2) гг. (по: Мартынов, 1983)
Fig. 72. The terms and hydrological
conditions of downstream
migration of Atlantic salmon smolts
in the Shugor River (after: Martynov,
1983)
ет продолжительным скатом
молоди в р. Печоре, когда на
их чешуе формируется прирост,
равный годовому. Кроме
того, встречаемость на
нерестилищах во второй половине
лета серебристых пе-
стряток (Никольский и др.,
1947; Владимирская, 1957;
Мартынов, 1979,1983; Сидоров, 1979а) может создать впечатление о длительном
пребывании покатной молоди в пресной воде. Мнение М.И. Владимирской
о продолжительном скате печорской молоди разделяет Д.К.
Дирин (1981), который основывает свои аргументы на анализе материалов
по пгуторской молоди лосося, представленных в работе В. П.Корниловой
(1967). На наш взгляд, расхождение в определении М.И. Владимирской
продолжительности речного периода жизни верхнепечорского
лосося по чешуе производителей и молоди связано с занижением возраста
последней по методическим причинам (см. гл. 3).
Наблюдения, проводившиеся нами за скатом молоди лосося на р. Щугор
в 60 км от устья реки в 1978 - 1979 гг., показали, что в этом притоке
Печоры скат проходит в довольно сжатые сроки (Мартынов, 1983). В 1978
г. продолжительность миграции покатников составила 12 дней (с 27 июня
по 8 июля), в 1979 г. - 14 дней (с 17 по 30 июня). При этом температура
воды колебалась от 8-9 °С в начале до 12-14 °С в конце ската (рис. 72). По
данным Г.П. Сидорова (1979а), покатники встречались в неводных уловах
при температуре воды 12-16 °С. Близкие значения температуры воды в
начале ската молоди атлантического лосося отмечаются в реках из других
районов Европейского Севера России (Сахончик, 1971; Бакштанский, Яко-
венко, 1976; и др.). Кратковременное понижение температуры воды до 5 -
6 °С в р. Щугор после начала миграции не приостанавливало её. Интенсивность
ската возрастала при подъеме воды. Окончание ската совпадало
с прогрессирующим падением уровня и возрастанием температуры воды.
Внешне покатники отличались от пестряток полной серебристой окраской
тела или в виде признаков посеребрения, когда просматриваются темные
поперечные полосы и красные пятна, свойственные речной окраске.
Таблица 84. Масса и длина покатников атлантического лосося р. Щугор в 1978 и 1979 гг.
(по: Мартынов, 1983 с изменениями)
Table 84. Weight and length of the Atlantic salmon smolts in the Shugor River in 1978 and
1979 (after: Martynov, 1983 with alterations)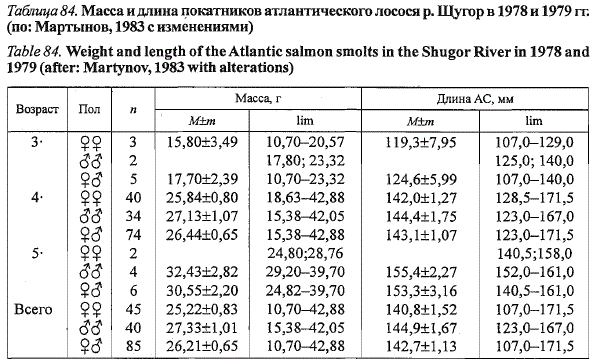
Средняя масса покатников щуторского лосося составила 26,2 г при
средней длине 142,7 мм (табл. 84). В уловах преобладали самки, составившие
56 % от всей молоди (учтены данные Г.П. Сидорова, 1979а). Среди
самцов обнаружено пять покатников с явно выраженными признаками
прошлогоднего нереста, что также отмечал Г.П. Сидоров. Гонады этих
самцов находились на II-VI стадиях развития, были увеличены в размерах
и выглядели воспаленными. На срезах гонад выделялась жидкость,
напоминавшая по цвету и консистенции сперму текучих самцов. Коэффициенты
зрелости бывших карликовых самцов (в среднем 0,42 % при
колебаниях 0,13-0,84 %) в четыре раза превышали коэффициенты зрелости
ювенильных самцов (в среднем 0,10 % при колебаниях 0,06 - 0,17 %)
и оказались близкими к коэффициентам зрелости самок. Самцы с признаками
прошлогоднего сперматогенеза составили 13,5 % от всех самцов,
или 6 % от выловленных покатников (и = 85).
Средняя продолжительность жизни покатников составила 4,01 года
(3— 5,9 %, 4- и 4+ - 87,1 %, 5— 7,0 %; п - 85 экз.) и совпала со средней
продолжительностью речного периода жизни щугорского лосося, определенной
по чешуе производителей (см. табл. 57). Интересно, что
среди молоди в уральских и тиманских притоках Печоры самки встречаются
в возрастных группах вплоть до 4+. Однако на Урале резкое
снижение доли самок наблюдается в возрастной группе пестряток 4+,
а на Тимане - 3+ (см. табл. 74), что свидетельствует о скате большинства
рыб в первом случае в возрасте четырех (4-), а во втором - трех
(3-) лет.
Только у одного покатника щугорского лосося на чешуе имелся прирост,
равный двум склеритам. Г.П. Сидоров (1979а) обнаружил приросты
в 1-2 склерита на чешуе девяти из 13 обследованных покатников. С учетом
сборов Г.П. Сидорова, рыбы с приростом на чешуе составили 10,3 %
от всей покатной молоди.
Совпадение продолжительности речного периода жизни щугорской
молоди, определенной по чешуе покатников и производителей, свидетельствует
о непродолжительном по времени скате смолтов по магистральному
руслу Печоры, а сходство в строении их чешуи говорит о незначительном
росте покатников в период ската. Таким образом, наши данные
не подтверждают представление о длительном (до года) скате молоди
лосося по руслу крупнейшей лососевой р. Печоры.
С другой стороны, по свидетельству местных жителей (д. Усть-Щу-
гор), молодь скатывается вниз по р. Щугор в августе-сентябре. По данным
О.В. Букова (1987), массовые миграции пестряток лосося наблюдаются
в летне-осенний период в бассейне Ильгча, притоке Печоры. При
облове мальковым неводом одних и тех же участков на притоках Печоры
- реках Щугор и Подчерем - ранней осенью (конец августа - начало
сентября) мы отмечали резкие колебания уловов старшевозрастных пестряток
в течение нескольких дней в одних и тех же местах (от нуля до 60 и
более рыбок на замет малькового невода). Эти колебания численности
могли быть связаны с миграцией молоди в реке.
Обсуждая вопрос об особенностях ската молоди атлантического лосося
в системах крупных рек, следует остановиться на рассмотрении временной
и пространственной динамики «миграционного окна», под которым
подразумевается относительно короткий промежуток времени, когда
условия среды благоприятны для совершения покатной миграции
(Saunders, Bailey, 1980). Один из главных факторов, характеризующих «миграционное
окно», - температура воды, в частности температурный по*
рог покатной миграции. Судя по наблюдениям за скатом молоди лосося в
р. Щугор, за температурный порог ската печорского лосося можно принять
температуру воды 10 °С. Вскрытие рек ото льда и прогрев воды в
бассейне Печоры происходят с юга на север (от верховий к низовьям) и с
запада на восток (от низовьев к верховьям в уральских притоках). Разница
между средними многолетними датами начала ледохода между верховьями
(р. Печора - с. Усть-Унья, 6 мая) и низовьями Печоры (р. Печора -
с. Оксино, 24 мая) достигает 18 дней, а в ее основном притоке р. Усе - 17
дней (ст. Сейда, 1 июня - с. Усть-Уса, 15 мая; Государственный водный
кадастр..., 1985). Только магистральное русло Печоры вскрывается ото льда
от верховьев к низовьям, а Усы - от низовьев к верховьям. Примерно такой
же интервал и географическая направленность должны иметь место
при прогреве воды до пороговых температур для ската молоди лосося.
Таким образом, в системе такой крупной реки, как Печора, «миграционное
окно» в магистральном русле перемещается по времени от верхо-
вий к низовьям, а в горных притоках от низовьев к верховьям. Можно
предположить, что осенние миграции пестряток в уральских притоках
Печоры являются адаптацией к особенностям динамики «миграционного
окна»: осенью молодь лосося из верховий гбрньтхрек концентрируется в
их нижнем течении, с тем чтобы весной оказаться в благоприятных для
ската условиях как в притоках, так и в магистральном русле. В результате
происходит предпокатная миграция пестряток печорского лосося, которые
смолтифицируются весной следующего года, быстро покидают притоки
и, попав в лабильное во времени и пространстве «миграционное окно»,
достаточно дружно достигают морских районов нагула в год ската. В магистральном
русле Печоры смолты лосося не зимуют. Крупные серебристые
пестрятки, которые появляются на нерестилищах в нижнем течении
притоков в конце лета (Владимирская, 1957; Мартынов, 1979), являются
десмолтами, т. е. бывшими покатниками, которые по тем или иным причинам
не «вписались» в «миграционное окно» и задержались в реке. Не
исключено, что серебристые пестрятки относятся к рыбам, не принимавшим
участие в предпокатной миграции и зимовавшим в верховьях горных
рек, где «миграционное окно» открывается в то время, когда в нижнем течении
притока и в магистральном русле Печоры orio закрывается. Крупная
серебристая молодь атлантического лосося известна и для других лососевых
рек (Черницкий, Лоенко, 1990; Мартынов, Куценко, 1985).
Скат молоди лосося в северных реках России в большинстве случаев
рассматривается щ основе наблюдений за ограниченное число лет. Для
оценки межпопуляционных особенностей покатной миграции молоди
представляют интерес данные о размахе межгодовой изменчивости ската
в той или иной реке, например многолетние наблюдения за скатом
молоди в реках Варзуга (Веселов, Калюжин, 2001) и Зимняя Золотица
(Мартынов, Дерец, 2005).
Средняя длина покатной молоди лосося р. Зимняя Золотица за ряд
лет составила 131,4 мм при средней массе 23,46 г (табл. 85) и низкой
межгодовой изменчивости. Самки и самцы не различаются по размерам.
Доля самок среди мигрантов в среднем составила 52 % (п = 325 экз.). За
десять лет наблюдений средняя продолжительность миграции покатни-
ков составила 29 дней (табл. 86). Самое раннее начало миграции отмечено
в конце мая 1977 г., наиболее позднее - 13 июня 1972 г. Миграция
завершалась в первой-второй декадах июля: 5 июля в 1975 г. и несколько
позже (13 июля) в 1978 г. Первые мигранты начинали встречаться в ловушке
при повышении температуры воды до 5,0 - 9,1 °С. По данным за
все годы исследований, основное количество рыб (54,5 %) скатывалось
при температуре воды от 15 до 18 °С (рис. 73, а). В различающиеся по
температурным условиям годы большинство мигрантов покидали реку
при разных значениях температуры воды (рис. 74).
В годы с теплой дружной весной (1972л 1974) более половины молоди
скатилось при температуре воды 17-19 °С, а в годы с затяжной
Табпща 85. Масса (P1, г) и длина (АС, мм) покатников атлантического лосося р. Зимняя
Золотица (по: Мартынов, Дерец, 2005)
Table 85. Weight (P,) and length (AС) of the Atlantic salmon smolts in the Zimnyaya Zolotitsa
River (after: Martynov, Derets, 2005)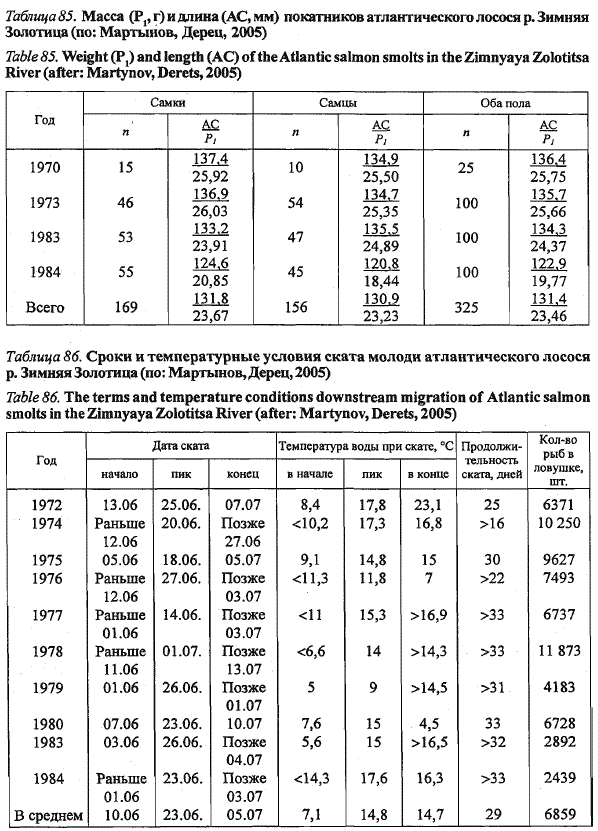
Таблица 86. Сроки и температурные условия ската молоди атлантического лосося
р. Зимняя Золотица (по: Мартынов, Дерец, 2005)
Table 86. The terms and temperature conditions downstream migration of Atlantic salmon
smolts in the Zimnyaya Zolotitsa River (after: Martynov, Derets, 2005)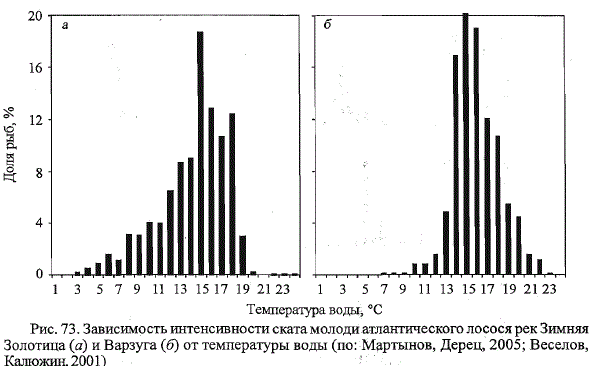
Рис. 73. Зависимость интенсивности ската молоди атлантического лосося рек Зимняя
Золотица (а) и Варзуга (б) от температуры воды (по: Мартынов, Дерец, 2005; Веселов,
Калюжин,2001)
Fig. 73. Dependence of migration intensivity of Atlantic salmon smolts in the Zimnayaya
Zolotitsa (a) and Varzuga (б) Rives on water temperature (after: Martynov, Derets, 2005; Veselov,
Kalyuzhin,2001)
весной при частом возврате холодов ( 1979) - при 9 °С и ниже. В разные
по условиям годы скат молоди проходил в разном диапазоне температур,
внутригодовые интервалы которых были существенно меньше,
чем при рассмотрении объединенных за ряд лет данных (см. рис.
73,д, 74). Часть молоди в 1978-1983 гг. скатывалась при температурах
воды, которые были значительно ниже пороговых при начале покат-
ной миграции. Например, в 1979 г. покатная миграция молоди началась
1 июня при температуре воды 5 °С (рис. 75). Затем в период с 5 по
12 июня температура воды снизилась до 2 °С. Данное похолодание прерывалось
однодневным повышением температуры воды до 8 °С. Первое
понижение температуры воды ниже пороговых значений сопровождалось
уменьшением интенсивности ската, но не привело к его остановке.
Лишь при повторном похолодании скат был приостановлен
на два дня.
Если в 1979 г. похолодание наступило в начале ската, когда температура
воды не превышала 6 °С, то в 1983 г. в середине ската, который начался
2 июня при температуре воды 5,6 °С и шел по нарастающей до 15 июня,
когда при температуре воды 11,3 °С в ловушку зашло 214 покатников. С 16
по 22 июня в результате возврата холодов среднесуточная температура
воды снизилась до 2,5-5,0 °С, т. е. до значений, ниже пороговых в начале
ската. Хотя скат молоди и продолжался весь недельный период похолодания,
но количество молоди в ловушке снизилось до 8-36 экз. в день.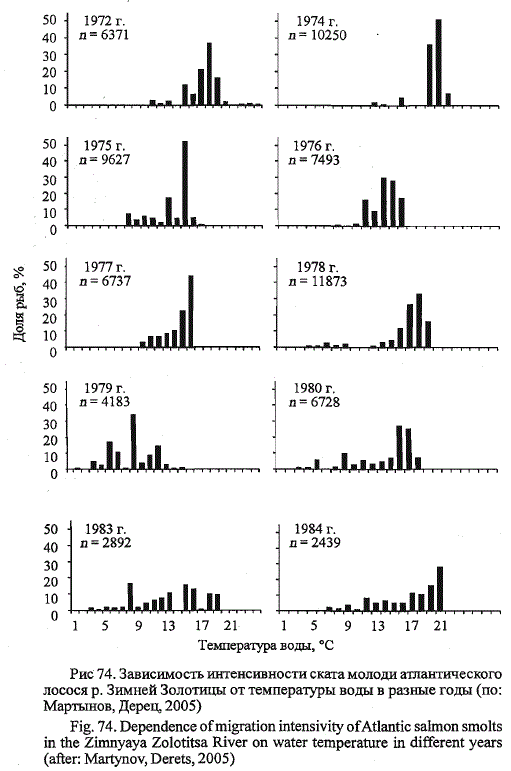
Рис 74. Зависимость интенсивности ската молоди атлантического
лосося р. Зимней Золотицы от температуры воды в разные годы (по:
Мартынов, Дерец, 2005)
Fig. 74. Dependence of migration intensivity of Atlantic salmon smolts
in the Zimnyaya Zolotitsa River on water temperature in different years
(after: Martynov, Derets, 2005)
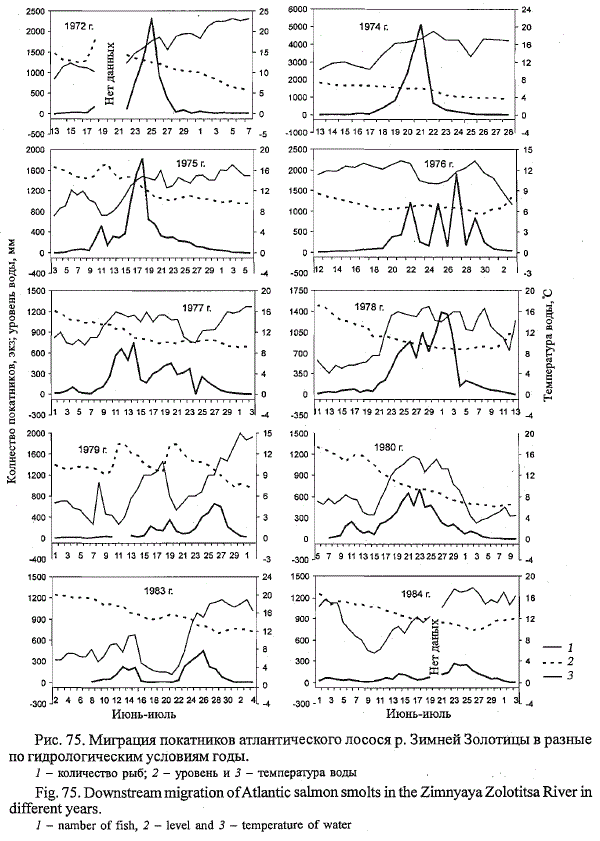
Рис. 75. Миграция покатников атлантического лосося р. Зимней Золотицы в разные
по гидрологическим условиям годы.
1 - количество рыб; 2 - уровень и 3 - температура воды
Fig. 75. Downstream migration of Atlantic salmon smolts in the Zimnyaya Zolotitsa River in
different years.
1 - namber of fish, 2 - level and 3 - temperature of water
Повышение 23 июня среднесуточной температуры воды до 7,5 °С вызвало
рост интенсивности ската молоди до 220 экз.
В 1978 и 1980 гг. при снижении температуры воды во время покатной
миграции ниже пороговых для начала ската интенсивность ската уменьшалась,
но скат не прекращался. Таким образом, можно констатировать,
что снижение температуры воды в период покатной миграции молоди
лосося в р. Зимняя Золотица до значений ниже пороговых при начале
ската не приводило к его остановке, но сильно снижало интенсивность
покатной миграции. В результате в годы с холодной затяжной весной наблюдалось
увеличение продолжительности ската в среднем на неделю
при формировании многовершинной кривой, отражающей его динамику.
Пороговые температуры покатной миграции молоди атлантического
лосося в р. Варзуге оказались на 2 °С выше (10-12 °С), а средняя продолжительность
покатной миграции - на 5 дней меньше (Веселов, Калю-
жин, 2001), чем в р. Зимняя Золотица. В последней значительная часть
молоди скатывалась при более низких температурах воды (см. рис. 73).
Вместе с тем по теплосодержанию воды сравниваемые реки не отличаются:
средняя многолетняя сумма градусо-дней воды в р. Зимняя Золотица
составила 1675,6+39,1, а в р. Варзуга - 1672±37,9 (см. Приложение,
табл. 1). На наш взгляд, наблюдаемые различия в покатной миграции
молоди лосося в рассматриваемых реках могут быть связаны с различиями
биологических характеристик рыб в популяциях. Так, покатная молодь
лосося в р. Зимняя Золотица значительно крупнее варзугской, что,
по-видимому, делает возможной миграцию части особей при более низкой
температуре воды.
Кривые, отражающие интенсивность ската молоди лосося в р. Зимняя
Золотица, можно подразделить на левую восходящую и правую нисходящую
части. До наступления пика миграции изменения ее интенсивности
под воздействием условий среды в меньшей мере ограничиваются таким
фактором, как величина контингента потенциальных мигрантов в реке. Данный
фактор становится доминирующим во второй фазе миграции, когда
основная часть молоди уже покинула реку, и именно он определяет выраженный
затухающий характер ската. Поэтому целесообразно рассматривать
связь интенсивности покатной миграции молоди лосося с гидрологическими
условиями раздельно для первой и второй фаз миграции (табл. 87).
Во все годы исследований, за исключением 1976-го, между интенсивностью
миграции и среднесуточной температурой воды на первой
фазе ската выявлена положительная зависимость (г = +0,51 ... +0,94). Ее
отсутствие в 1976 г. объясняется началом миграции молоди при высокой
пороговой температуре и дальнейшим продолжением миграции при незначительных
колебаниях температуры воды (см. рис. 75). В данном случае
динамика миграции, по-видимому, слабо модифицировалась условиями
среды и в основном определялась факторами внутреннего порядка,
связанными с физиологической готовностью рыб к скату.
198
Таблица 87. Коэффициенты корреляции между количеством покатников атлантического
лосося р. Зимняя Золотица и факторами внешней среды (по: Мартынов, Дерец, 2005)
Table 87. Correlation coefficients between downstream migration intensivity of Atlantic salmon
smolts in the Zimnyaya Zolotitsa River and environmental factors (after: Martynov, Derets,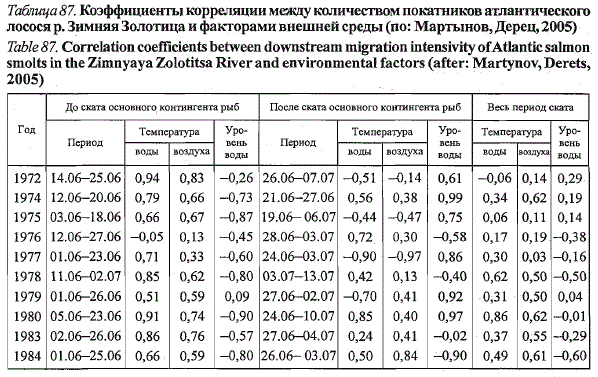
Во второй, нисходящей, фазе покатной миграции связь между интенсивностью
ската и температурой воды в разные годы была противоречивой:
от высокой положительной до столь же высокой отрицательной (см.
табл. 87). Данное обстоятельство, на наш взгляд, свидетельствует о случайности
рассматриваемой зависимости. Высокая межгодовая изменчивость
связи определялась затухающим характером самой покатной миграции
на заключительном этапе, с одной стороны, и противофазными
межгодовыми колебаниями погодных условий (от прогрессирующего
прогрева воды до ее сильного выхолаживания при позднем возврате холодов)
- с другой.
Связь между интенсивностью ската молоди и температурой воды за
весь период покатной миграции в большинстве случаев оказалась незначимой
(см. табл. 87). Высокие положительные коэффициенты корреляции
между сравниваемыми параметрами в 1978 и 1980 гг. определяются
совпадением затухающего ската во второй фазе с понижением температуры
воды при возврате холодов.
Аналогичные результаты были получены при изучении связи интенсивности
ската молоди с температурой воздуха с той лишь разницей, что
значения коэффициентов корреляции были несколько ниже из-за известной
инертности прогрева и охлаждения воды по сравнению с соответствующими
изменениями температуры воздуха. Таким образом, температурные
условия оказывают выраженное влияние на динамику покатной
миграции молоди лосося в р. Зимняя Золотица в ее первую фазу, т. е.
до ската основного контингента мигрантов. Во вторую фазу покатной
199
миграции изменения температуры среды также оказывают определенное
модифицирующее воздействие на скат, но в меньшей степени.
Анализ зависимости между интенсивностью ската молоди и уровен-
ным режимом воды показал, что в первой фазе покатной миграции в большинстве
случаев наблюдается высокая отрицательная, а во второй — положительная
связь (см. табл. 87). Выявленные зависимости, на наш взгляд,
носят формальный характер и определяются тем, что покатная миграция,
как правило, проходит на фоне прогрессивно снижающегося уровня воды.
В первой половине динамика интенсивности ската молоди находится в
противофазе с динамикой уровня воды, а во второй совпадает с ней. При
сопоставлении интенсивности ската молоди с уровнем воды за весь период
покатной миграции происходит взаимное гашение отрицательно и
положительно значимых связей первой и второй фаз миграции, и мы не
обнаруживаем зависимости между сравниваемыми параметрами, что
совпадает с действительным положением вещей.
Покатная миграция молоди атлантического лосося связана со сменой
среды обитания и имеет внутреннюю мотивацию, которая определяется
физиологической готовностью рыб к переходу из гипотонической среды
в гипертоническую. Рассмотренная выше межгодовая изменчивость динамики
покатной миграции молоди лосося р. Зимняя Золотица является
результатом взаимодействия внутренних факторов, определяющих степень
готовности рыб к скату, и условий среды, модифицирующих как степень
готовности рыб к скату, так и динамику покатной миграции. В ряде
публикаций показано, что покатная миграция молоди лосося начинается
при весеннем прогреве воды до определенной суммы градусо-дней, которую
можно рассматривать как пороговую величину, запускающую по-
катную миграцию (Бакштанский и др., 1976; Нестеров, 1985). В этом случае
становятся понятными межгодовые различия среднесуточной температуры
воды в начале ската. Пороговые значения суммы градусо-дней
могут достигаться как при медленном прогреве, когда среднесуточные
температуры воды имеют низкие показатели, так и при быстром прогрессирующем
прогреве, когда среднесуточная температура к началу ската
достигает относительно высоких значений.
По данным А.Е. Веселова и СМ. Калюжина (2001), зависимость покатной
миграции молоди варзугского лосося от накопленной суммы тепла
не выявлена, однако приводимые авторами суммы градусо-дней воды
до начала покатной миграции молоди лосося достаточно близки в разные
по гидрологическим условиям годы. Косвенным указанием на наличие
в этой реке зависимости начала покатной миграции молоди от порогового
значения суммы тепла может служить межгодовая изменчивость
среднесуточной температуры воды при начале покатной миграции, а также
отмеченный авторами случай начала покатной миграции варзугской
молоди лишь при повторном прогреве воды до среднесуточного порогового
значения.
200
На Севере России температурные условия в весенний период весьма
непостоянны по годам, что отражается на динамике покатной миграции
молоди лосося, определяя ее высокую межгодовую изменчивость. В дружные
теплые весны большинство рыб скатывается в море в течение 3-4
дней при высоких значениях среднесуточных температур воды. В холодные
весны при частом возврате холодов интенсивность ската молоди в
пределах миграционного периода выравнивается, а скат рыб проходит
при более низких температурах. Температурная модификация сроков и
интенсивности покатной миграции молоди лосося может иметь адаптивное
значение для соответствующей межгодовой температурно-зави-
симой изменчивости трофических условий ската в рамках так называемого
«миграционного окна».
Смотрите также