Биологическая структура популяций кижуча
Возрастной состав кижуча
у азиатского кижуча встречаются рыбы 11 возрастных категорий, у североамериканского - 9 (табл. 81), а теоретически, сучетом ската молоди в море в возрасте 0+.4+ и возврата на нерест как в годската, таки через 1-2 морских года жизни возрастной состав кижуча как вида может быть представлен 14 группами.
Таблица 81. Встречаемость возрастных группировок кижуча в нерестовых стадах
Основными возрастными категориями у кижуча материкового побережья Охотского моря, которые ежегодно встречаются в подходах и формируют около 99% возвратов, являются 1.1, 2.1 и g
За весь период наблюдений в североохотоморских реках кижуч в возрасте 2.1 формировал около 58,7% подходов, а на долю рыб в возрасте 1.1 приходилось около 37,8%. В реках Охотского района, наоборот, подходы главным обраBOM COCTORITM H3 ಸಿ? возрасте 1.1 — 69,6%, а доля рыб в возрасте 2.1 была около 30,1%. Что касается доли рыб в возрасте 3.1, то в реках Охотского района за весь период исследований она составила около 0,3%, а в североохотоморских реках доля таких рыб в подходах была на порядок выше — 3,5%.
Небольшую часть в подходах охотоморского кижуча составляет такая редкая возрастная группа, как каюрки. На долю таких рыб в подходах приходится до 1%. в возрасте 1.0 — 0,3%, в возрасте 20 — 0,3—1,0%,
В.И. Грибанов (1948) считал, что каюркивстречаются у кижуча во всех камчатских реках. На юго-западе Камчатки в оз. Б. Вилюй доля каюрок колебалась от 10,8 до 21,9% (Мельникова и др., 2004). Такая высокая численность каюрок в этой популяции объясняется благоприятными условиями обитания молоди в озере. На одном из северных Курильских островов (о. Шумшу) в р. Беттобу найдено 8,7% каюрок. Однако в других водоемах каюрок не было обнаружено (Гриценко и др., 2000; Стыгар и др., 2000). На Сахалине каюрки встречаются исключительно редко (Гриценко, 2002).
На североамериканском побережье какорок называют джеками — «jacks». В р. Колумбия (Британская Колумбия) отмечено около 6% джеков (Магг, 1943),
в р. Ковичан — до 13% (Neave, 1949), в руч. Минтер (Вашингтон) их возврат колебался от 21 до 27% (Salo, Bayliff, 1958), в оз. Тэн Майлз (Орегон) их доля достигала 46% (Morgan, Henry, 1959), а в руч. Свельтзер (Британская Колумбия) возврат джеков был выше (до 74%), чем крупных проходных рыб (Foerster, Ricker, 1953). В целом можно заметить, что на североамериканском побережье относительная численность скороспелых мелких самцов кижуча гораздо выше, чем на азиатском. Феномен «карликовых» самцов кижуча X. Билтон с соавторами (Bilton et al., 1984) связывают с более ранними, чем обычно, сроками скатаболее крупной молоди, что приводит к увеличению доли джеков в возвратах.
Рыбы, проведшие 2 года в море (1.2), в подходах исследованных популяций кижуча составляют небольшую часть (0,4—0,5%), рыбы возрастной группы 2.2 — 0,4—3,4%. Невелика их доля и в других водоемах. Например, в реках Восточной Чукотки они составляют около 5,5% подходов (Черешнев, Агапов, 1992а), на Северных Курилах — 2,7% (Гриценко и др., 2000). Единично рыбы в возрасте 1.2 и 2.2 встречаются и на Камчатке (Зорбиди, 1970б). Увеличение доли особей кижуча с двумя пресноводными годами отмечено в водах Берингова моря и Тихого океана, прилегающих к Восточной Камчатке (Зорбиди, 2003).
У кижуча в динамике соотношения возрастных групп время от времени наблюдается смена доминантной возрастной категории. Например, попеременное доминирование кижуча в возрасте 1.1 и 2.1 наблюдается на Сахалине (Чуриков, 1975; Ковтун, 1994). Не является исключением и кижуч материкового побережья Охотского моря, у которого смена доминирующей возрастной категории наблюдалась неоднократно. Например, у североохотоморского кижуча рыбы, прожившие в реках 1 год, составляли основу подходов во второй половине 1960-х гг., а также в период со второй половины 1970-х по первую половину 1990-х гг., а в остальные периоды в нерестовом стаде преобладали особис 2 речными годами жизни. В реках Охотского района рыбы с 1 речным годом жизни доминировали в подходах во второй половине 1960-х — первой половине 1970-х гг., а также в 1980-х — первой половине 1990-х гг. и в 2000-е гг. Во второй половине 1970-х гг. доля рыб с 1 и с 2 речными годами была практически одинаковой. И только в первой половине 1960-х и второй половине 1990-х гг. в его подходах преобладали рыбы с 2 речными годами жизни (рис. 63).
Доля рыб с 3 речными годами всегда была низкой, но в североохотоморских реках в отдельные годы они составляли до 9,7—22,2% подходов, а с 1990-х гг. по настоящее время их количество в подходах в реки северного побережья увеличилось с 1,0 до 6,9%. Примечательно, что доля рыб с 3 речными годами, как правило, возрастала в годы высокой численности рыб, проведших в реках 2 года (см. рис. 63).
Длина и масса тела кижуча
Кижуч — третий по размерам тела вид тихоокеанских лососей. Подлине и массе он уступает только чавыче и кете.
В североохотоморские реки заходит кижуч, имеющий длину от 27,0 до 82,1 см и массу тела от 0,29 до 8,55 кг. Средние значения данных признаков составляли, соответственно, 65,5 см и 3,79 кг.
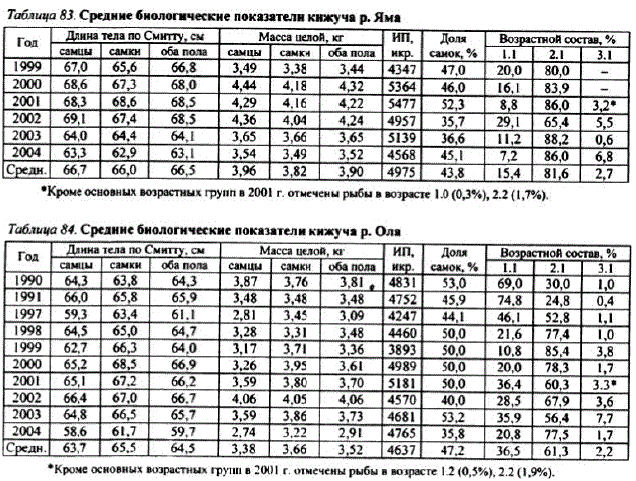
Распределение кижуча подлине — мономодальное и имеет нормальное распределение как в целом по побережью, так и по рекам в отдельности. Основная масса рыб имеет длину от 58 до 75 см, а вариационный ряд самцов немного шире в сравнении с таковым у самок, так как самцы имеют большую изменчивость по этому показателю, в частности, каюрки у североохотоморского кижуча представлены исключительно самцами (рис. 64).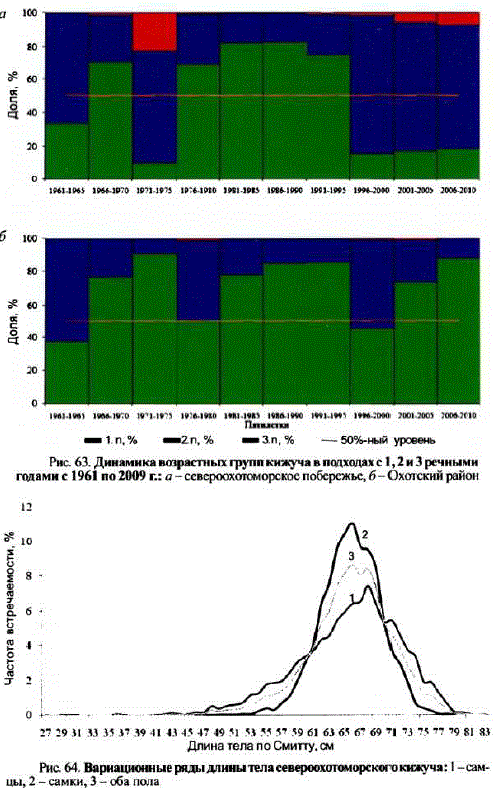
Масса каюрок (скороспелых рыб, проведших 1—3 года в реке и однолето в море) колебалась от 290 до 850 г, длина — от 27 до 38 см. Встречаются каюрки редко, обычно во второй половине — конце нерестового хода. Анадромную миграцию совершают вместе с крупными производителями. Размеры северокурильских каюрок колебались в пределах 33—50, в среднем 39 см, колебания массы тела составили 440—630 г (Стыгар и др., 2000). Средняя длина джеков из водоемов Британской Колумбии варьировала от 30 до 35,6 см (Foerster, Ricker, 1953;Anderson, Narver, 1975; Fraser et al., 1983), для джеков из Калифорнии она cocTaBHIJIa 40,9 cM (Shapovalov, Taft, 1954).
Полинейно-весовым показателям рыбы младших возрастов всегда мельче старшевозрастных рыб (табл. 82). И эти различия прослеживаются как по годам возвратов, так и по поколениям. Интересно, что в возрастной категории 1.1 самки всегда крупнее самцов. Вероятно, это приспособительная реакция, позволяющая увеличить репродуктивные возможности, поскольку известно, что умногих рыб более крупные особи имеют более высокую плодовитость или более крупную икру. Исходя из того, что в возрасте 1.1 самки крупнее самцов, а в остальных возрастных группах они мельче, нельзя говорить о половомдиморфизме у североохотоморского кижуча без разделения рыб по возрасту.
Таблица 82. Изменчивость линейно-весовых показателей североохотоморского кижуча B 2001 —2009 TT,
Возраст, лет

Характеристика показателей биологической структуры основных популяций кижуча приводится в табл. 83—86. Размерно-весовые показатели кижуча материкового побережья Охотского моря отличаются по отдельным годами районам воспроизводства. Наибольшую длину тела имеет кижуч рр. Яма и Тауй — среднемноголетние показатели этого признака составляют соответственно, б6,5 и 66,1 см. Наибольшей средней массой тела отличается кижуч р. Тауй — 4,25 кг. Наибольшей средней плодовитостью выделяется кижуч р. Яма — 4975 икр,
В течение нерестового хода обычно наблюдается снижение размерновесовых показателей кижуча к концу миграции (табл. 87). В некоторые годы по одним рекам подобные изменения имеются, по другим такой закономерности не наблюдается.
Кроме того, на материковом побережье Охотского моря прослеживается клинальная изменчивость размерно-весовых показателей кижуча: размер и масса тела рыбувеличиваются с северо-востока на юго-запад (табл. 88).
У кижуча Азиатского и Североамериканского континентов значительных различий по средним показателям длины и массы тела не наблюдается, на Дальнем Востоке наиболее крупными размерами выделяется кижуч из р. Тымь северного Сахалина (табл. 89).
Таблица 87. Изменение средних размерно-весовых показателей кижуча в течение нерестовой миграции
2002r, 2003, Номер р. Ола p. Tayl р, Ола p, Tay пробы (дата) | длина, масса, | длина, масса, длина, масса, длина, 
Плодовитость кижуча
Средняя индивидуальная плодовитость североохотоморского кижуча составляет 4645 икр. Она варьировала в очень широких пределах — от 1180 до 11 160 икр., и основная доля самок (около 85%) имела плодовитость от 3300 до 5900 икр. Распределение вариационного ряда индивидуальной плодовитости кижуча близко к нормальному (рис. 65).
Рис. 65. Вариационный ряд индивидуальной плодовитости североохотоморского кижуча
Плодовитость, а также масса икринки у кижучанс оставались постоянными величинами. В их динамике наблюдались периоды как снижения, так и роста. Например, с 2004 по 2008 г. масса икринки упала до 1 г. х 10 и ниже в то время как плодовитость достигла своих максимальных значений — это был период снижения размеров североохотоморского кижуча. В целом в первом десятилетии XXв, его индивидуальная плодовитостъ существенно снизилась: с 4896— 5275 икр, в начале десятилетия до 3958—4261 к его окончанию,
Как показано на рис. 66, плодовитость (и индивидуальная, и относительная) и масса икринок изменялись в противофазе, корреляция между индивидуальной плодовитостью и массой икринки была -0,55, а между относительной плодовитостью и массой икринки -0,90.
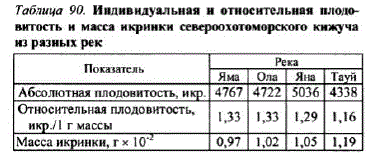
Из рек североохотоморского побережья, имеющих наибольшие запасы кижуча, — Яма, Ола, Яна и Тауй, наименьшей средней плодовитостью характеризовался кижуч р. Тауй — 4338 икр., наибольшей — кижуч р. Яна — 5036 икр. В географической изменчивости этого показателя клина не прослеживалась, но наблюдалось увеличение массы икринки и снижение относительной плодовитоcTH (TaŐJI. 90).
Казалось бы, имея крупные размеры и относительно низкую плодовитость, кижуч р. Тауй должен хуже остальных исследованных популяций поддерживать свою численность, отвечая на изменения лимитирующих факторов среды. Однако его запасы в этой реке одни из самых высоких. В данном случае, очевидно, приспособление к поддержанию численности пошло по пути увеличения MCCLII MKpMHKM.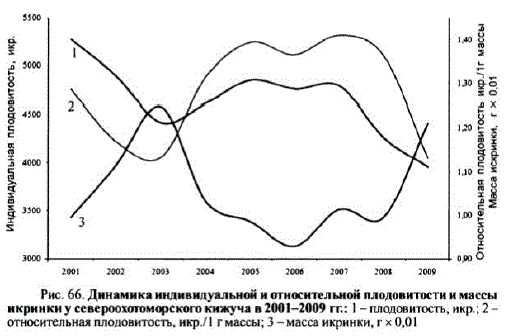
Рис. бб. Динамика индивидуальной и относительной плодовитости и массы икринки у североохотоморского кижуча в 2001—2009 гг.: 1 — плодовитость, икр.; 2 относительная плодовитостъ, икр./1 гмассы; 3 — масса икринки, гx 0,01
Как справедливо отмечал Г.В. Никольский (1974), при изменении плодовитости изменяется и размерикринок, что определяет продолжительность питания за счет желтка, размеры личинки при переходе на внешнее питание, а значит, и качество необходимых для личинки кормов на первых этапах личиночного периода. Таким образом, тауйский кижуч запасает в икре наибольшее количество питательных веществ, что, несомненно, способствует его выживаемости.
Прямыми наблюдениями за размерами личинок тауйского кижуча мы не располагаем, однако есть работа Н.И. Виленской (2002) по другому тихоокеанскому лососю — чавыче, в которой показано, что из крупной икры чавычи
Taf 9). выклевываются крупсIIJлыЧет ндивидуальная и относительная плодовитость и масса икринки североохотоморского кижуча рые раньше встают на из разных рек но, раньше переходят Яма Ола Яна Тауй, Абсолютная плодовитость, чем молодь, выклюнувшаяся из мелкой икры уляцcкого кижуча, вероятно, развивалась по другому пути поддержания высокой численности — он имеет высокую относительную плодовитость и самую низкую из исследованных популяций массуикpMHoK (Ta6JI. 9l).
Относительная IJIOLOBHTOCTb. HBUIHeTC OKa3aTeleM физиологической CIUсобности самок продуцировать определенное количество икры на единицумассы и зависит от условий ростарыбинагула (Решетников, 1980). Оценить условия нагула по нашим данным не представляется возможным, однако просле
дить изменчивость признака по возрасту и массе вполне реально,
Из исследованных популяций только у кижучар. Тауй с увеличением возраста наблюдался рост средней плодовитости. У кижучар. Яма наибольшую
Показатель
Плодовитость имели рыбы в возрасте 2.1, а У ЯНСКОГО И ОЛЬСКОГО кижуча рыбы в этом
же возрасте имели наименьшую плодовитость. Более информативной оказалась отнотость кижуча, которая в ямской, ольской и янской популяциях с воз тауйской имела слабую тенденцию к увеличена разнонаправленныи характер измене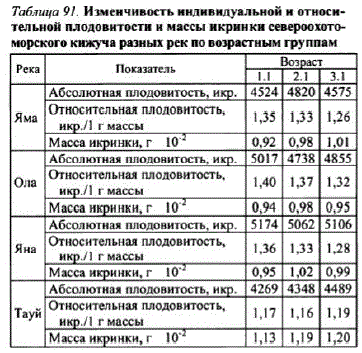
Разнонаправленность изменений плодовитости (как индивидуальной, так и относительной) и массы икринок по возрастам, по нашему мнению, свидетельствует о том, что возраст не является ключевой причиной изменчивости этих показателей. Определяющей оказалась зависимость плодовитости от массы тела (рис. 67, a). Для получения четкой картины были отобраны только массовые классовые интервалы, содержавшие не менее 50 особей. Уровень корреляции между плодовитостью и массой тела составил для ямского кижуча 0,99, для ольского — 0,98, для янского — 0,95 и для тауйского — 0,93. Уровень значимости во всех случаях превышал 0,001.
Наряду с увеличением индивидуальной плодовитости наблюдалось снижение относительной плодовитости (рис. 67, б) и, соответственно, ростмассы икринок (рис. 67, в). Исключение составил кижуч р. Яма, у которого наблюдалось незначительное снижение относительной плодовитости, а масса икринки практически неизменялась.
По-видимому, при выполнении рыбоводных мероприятий по воспроизводству запасов североохотоморского кижуча следует отбирать крупных производителей, которые хотя и имеют более низкую относительную плодовитость, но производят более крупную икру, что должно положительно сказаться на качеCTBC MOJIOJILM M 202 B bIXKIM BaCMOCTIH.
На Камчатке, где имеются самые большие запасы кижуча, колебания плодовитости составляют 2800—7100 икр. (Смирнов А.И., 1960), его средняя плодовитость по годам варьирует от 3671 до 5343, а среднемноголетние показатели по двум основным рекам составили 4534 (р. Камчатка) и 4492 (р. Большая) икр. (Зорбиди, 1970б, 1975). Сходные средние величины плодовитости для Камчатки приводят и другие исследователи (Грибанов, 1948; Грачев, 1968).
Отметим, HTO IIOKa3aTCJIH IIJIOIOBHTOCTH охотоморского KMXKy"Ha выше, чем камчатского. Размах плодовитости сахалинского кижуча составляет 1760—9010, в среднем 4320-5370 икр (Гриценко, 1973, 2002).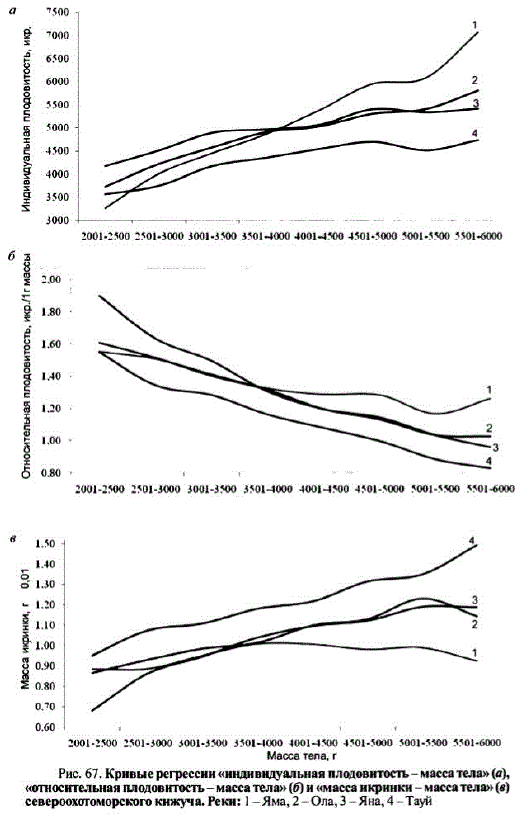
Маоса тела, г Рис. 67. Кривые регрессии «индивидуальная плодовитостъ— массатела» (а), «относительная плодовитость — масса тела» (б) и «масса икринки — массатела» (в) североохотоморского кижyча. Реки: 1 - Яма, 2 - Ола, 3 - Яна, 4-Тауй
Северокурильский кижуч имеет колебания плодовитости 3045—6160, в среднем 4722 икр. (Стыгар и др., 2000). Колебания плодовитости чукотского кижуча составляют 2394—5728, в среднем 3430—4993 икр. (Черешнев, Агапов, 1992а). Наибольшей плодовитостью среди азиатского кижуча выделяется популяция р. Авьяваям (Камчатка) — его средняя плодовитость 6992—7079 икр. (Грачев, 1968). В целом плодовитостъ охотоморского кижуча приближается к плодовитости позднего камчатского кижуча (Грибанов, 1948; Зорбиди, 1970б) и сахалинского (Гриценко, 1973). Относительно высокая средняя плодовитость кижуча при сходных с другими видами рода размерами обусловлена меньшим диаметром икринок.
Абсолютная плодовитость североамериканского кижуча значительно ниже как по максимальным, так и по средним показателям. Большей плодовитостью характеризуются популяции кижуча Аляски: оз. Карлук — средняя 4706 (колебания 1724—6906) икр. (Drucker, 1972), р. Свэнсон — среднее 3149 4023 (Engel, 1967). Средняя плодовитость кижуча Британской Колумбии из р. Найл составJIAer 23.1.0 (Wickett, 1951), H3 p. Bhr KBOJIMKyM - 2574 (Fraser et al., 1983) HKp. B штатах Вашингтон и Орегон она еще ниже — 2500 (Salo, Bayliff, 1958) и 1983 (Koski, 1966) икр. соответственно.
Упитанность, зрелость
Упитанность самцов кижуча несколько выше, чем самок, и коэффициент их упитанности колеблется от 0,62 до 2,17; в среднем по отдельным популяциям он варьирует в пределах 1,25—1,70. Индекс упитанности самок меняется от 0,96 до 2,44; в среднем для разных популяций ее диапазон составляет 1,21—1,61. Упитаннось самцов несколько выше — в среднем изменялась от 1,25 до 1,70 (табл. 92). Упитанность, как праBMJIO, IIOHMKaCTCH KKOHLly HepecTOBOTO Xosa,
Таблица 92. Коэффициенты зрелости и упитанности охотоморского кижуча
что объясняется уве- Гонадо-соматичcский Коэффициент JIHHCHCM LOJIM MCVIK IX, Река индекс, % упитанности по Кларк более молодых рыб и ML MHH CMLL i 
товой миграции проис
ходят изменения полового состава. В начале хода отмечается преобладание (до 60—65%) самцов, затем к середине хода соотношение самцов и самок выравнивается, в конце хода обычно преобладают самки — до 70%, т. е. структура и динамика нерестовой миграции у кижуча такая же, как и у других видов лососей.
Гонадо-соматический индекс (ГСИ) (отношение массы гонад к массе поротой рыбы) у самцов кижуча варьирует от 4,09 до 15,71, его средние показатели для основных рек изменялись от 7,74 до 10,35. Амплитуда изменчивости этого признака у самок еще больше — 5,63—27,16, а его средние значения колебались от 15,04 до 17,10 (см. табл. 92).
Отметим, что индекс зрелости выше у рыб, заходящих в реки небольшой протяженности (pp. Сиглан, Мотыклейка) и меньше для рек с большой длиной водотока (pp. Яма, Тауй). ГСИ самок увеличивается к концу анадромной миграции. С приближением к нерестилищам он увеличивается до 26—28, а масса гонад может достигать 1300 г. Увеличение массы гонаду самок происходит за счет иммобилизации каротиноидов и энергетических веществ из мышц.
У самцов наибольшие значения ГСИ наблюдаются, как правило, в начале — середине нерестового хода. Это связано с тем, что в отличие от самок у самцов в течение нерестовой миграции не происходит значительного увеличения массы половых продуктов. Высокая массагонад в первой половине хода у самцов обусловлена накоплением энергетических веществ, которые в дальнейшем идут на формирование и развитие сперматоцитов и спермиев. Период относительно быстрого сперматогенезаулососей происходит после перехода из воды морской солености в пресную, затем наступает этап более медленного развития гонад. На нерестилищах в период размножения продуцирование и эйякуляция спермиев у них может продолжаться до 3—4 недель (Смирнов А.И., 1975). Один самец может принимать участие в нересте с несколькими самками.