Особенности воспроизводства североохотских лососей и их сравнительная характеристика
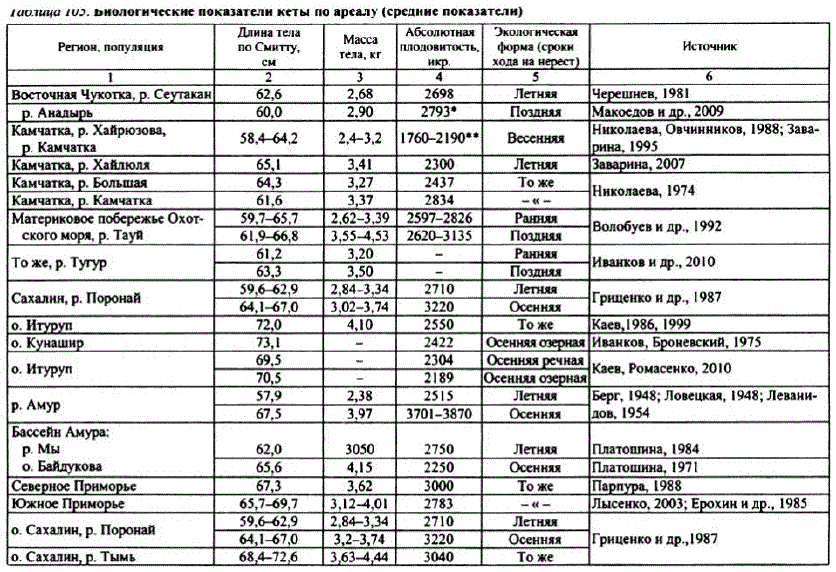
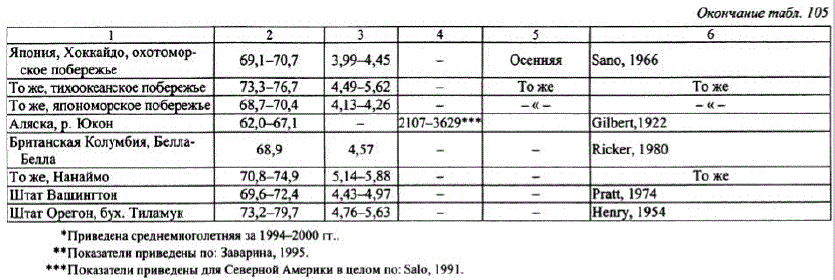
Кета. Со времени описания Л.С. Бергом (1948) сезонных раскеты в бассейне Амура не прекращаются публикации о расах и экологических формах кеты по азиатской части ареала (Абрамов, 1948; Бирман, 1964; Иванков, 1972; Волобуев, 1984, Гриценко и др., 1987, Николаева, Овчинников, 1988. Заварина, 1995; Иванков, Броневский, 1975; Махинов, Золотухин, 1999; Кузищин и др., 2010; Иванков и др., 2010; и др.). Существующие расы и формы кеты на Дальнем Востоке выделяются на основе следующих дифференцирующих признаков: сроков нерестового хода, особенностей биологии и репродуктивной экологии. По срокам анадромной миграции дальневосточная кета подразделяется на весеннюю (майиюнь), летнюю (июль-август) и осеннюю (сентябрь). Наибольшего внутривидового разнообразия кета достигает на Камчатке. Здесь описаны весенняя, летняя ранняя, летняя поздняя и осенняя формы кеты, различающиеся по срокамнерестового хода и нереста, распределением и топографией нерестовых гнезд в бассейнах нерестовых рек в зависимости от их геоморфологии и гидрологического режима (Николаева, Овчинников, 1988; Заварина, 1995, 2007; Кузищин и др., 2010). Причем и летняя, и осенняя формы могут размножаться как на участках рек с подрусловым водоснабжением, так и на выходах грунтовых вод, образуя русловой и ключевой экотипы (Кузищин и др., 2010).
По особенностям воспроизводства кета отличается предпочтением к биотопам с разным типом водоснабжения. Если раньше клетней кете относили особей вида, размножающихся в летнее время на участках рек с подрусловым водоснабжением, а к осенней расе — рыб позднего осеннегохода на нерест с воспроизводством на выходах грунтовых вод, то сейчас эти признаки, маркировавшие расы, довольно условны. Описан ряд форм кеты, различающихся сроками нерестового хода, местами и биотопами размножения. Например, кета Камчатки весеннего и летнего сроков хода на нерест размножается включах и лимнокренах на выходах грунтовых вод (Крохин, Крогиус, 1937; Бирман, 1964; Смирнов А.И., 1975). Однако часть кеты летнего хода воспроизводится на участках рек с подрусловым водоснабжением(Леман, 1992, Кузищин и др., 2010). Кета позднегохода может размножаться вместах, не имеющих выходов грунтовых вод (Абрамов, 1948). По данным К.В. Кузищина с соавторами (2010), камчатская осенняя кета размножается на мощных выходах ключей вместах апвеллингов грунтовых вод, а ранняя летняя — на подрусловом потоке, вместах инфильтрации руслового потока валлювий.
В то же время в бассейне Амура установлено четкое разграничение нерестующей кеты на летнюю, воспроизводящуюся наводах подруслового потока, иногда с примесью более теплых глубинных вод, и осеннюю, откладывающую икру на выходах грунтовых вод включах, лимнокренах (Кузнецов И.И., 1937, Леванидов, 1969; Смирнов А.И., 1975). Однако, несмотря на устоявшееся мнение относительно существования в бассейне Амураи в реках Сахалинакеты указанных рас, в бассейне Амура (р. Амгунь) и на Сахалине (бас. р. Поронай) описан экотип осенней кеты, воспроизводящийся на подрусловом потоке (Золотухин, 2009).
То есть существует довольно пестрая картина репродуктивных предпочтений рас и форм кеты по азиатскому участку ареала. Разнообразие жизненных стратегий и особенностей репродуктивной экологии, очевидно, следует рассматpивать как результат взаимодействия генотипа с условиями среды. А экологическое разнообразие способствует более широкому освоению ареала, многообразию и процветанию вида.
Все внутривидовое разнообразие выявленных к настоящему времени темпоральных, сезонных и экологических (экотипы) форм кеты, очевидно, не следует сводить к двум генерализованным условным категориям: «русловой» и «ключевой», как предлагают В.Н. Иванков с соавторами (2010). Внутривидовая диверсификация кеты зависит от разнообразия факторов среды обитания, и как вид, экологически пластичный, она способна образовывать «букеты» или «пучки» жизненных форм, которые с успехом можно использовать в практической деятельности, например, в рыбоводстве, аквакультуре, организации рекреационного рыболовства. При этом надо будет руководствоваться сроками нерестового хода, генетическими характеристиками, учитывать особенности биологической структуры и топографию нерестилищ, присущие каждой из выделенных хорологических и биотопических группировок. В связи с этим считаем, что нет убедительных оснований для такого узкого, упрощенного подхода к оценке биологического разнообразия полиморфного, экологически поливалентного вида, каким является кета, В реки материкового побережья Охотского моря заходит на нерест кета двух сезонных и экологических групп. По срокам нерестовой миграции выделяСтся кета раннего летнего (июнь-июль) и позднего летне-осеннего (август-сентябрь) хода. Кета выделенных форм может воспроизводиться как симпатрично в бассейне одной реки, так и раздельно, когдата или иная форма для размножения выбирает разные реки. По особенностям репродуктивной экологии охотоморская кета является аналогом летней и осенней расамурской кеты в классическом понимании (Берг, 1948) как по основным биологическим показателям, таки при выборе мест размножения. Ранняя кета мельче поздней, нерестится на участках рек с подрусловым водоснабжением. Поздняя кета крупнее и размножается на выходах грунтовых вод. Кета указанных форм обитает на всем протяжении материкового побережья Охотского моря — отрек зал. Шелихова на северо-востоке до зал. Тугурский на юго-западе (Волобуев и др., 1990, Кульбачный, Иванков, 2011),
Очевидно, наличие на участке нерестового ареала азиатской кеты разнообразных условий позволяет воспроизводиться определенным экологическим модификациям кеты, реализованным в виде той или иной формы жизненных стратегий. Они наиболее приспособлены к условиям среды или же, заселив исторически данный район обитания, являются экологически более адаптированными по отношению к другим сезонным формам и более конкурентоспособными для того, чтобы оставаться доминирующими.
По биологическим показателям ранняя охотоморская кета ближе всего стоит к камчатской весенней форме, летней амурской и сахалинской, а поздняя — к камчатской летней (табл. 105).
В размножении охотоморской кеты доминирует естественное воспроизводство. Возвраты кеты от выпусков заводской молодине превышают 11% от численности ее общих подходов. Оценка численности производителей охотоморской кеты на нерестилищах проводится с 1966 г. в двух основных районах: Североохотоморском и Охотском. В Аяно-Майском и Тугypo-Чумиканском районах авианаблюдения за их численностью систематически не проводились.
Пропуск на нерестилища кеты в реки Североохотоморского района (в пределах Магаданской области) в отдельные годы колебался от 182 (1998 г.) до 2977 (1966 г.) тыс. рыб.
В Охотском районе Хабаровского края наименьшее количество производителей кеты отмечено в 1973 г. — 159 тыс. рыб, максимальное — в 2002 г. — 4386 тыс. рыб. За последние 15 лет средняя численность пропущенных на нерест производителей кеты по Северному побережью составила 1098, по Охотскому району — 2898 тыс. рыб. Максимальные пропуски кеты на нерест по побережью (5769 и 5424 тыс. рыб) были в 2006 и 2009 г. соответственно,
Удельный вес основных рек материкового побережья Охотского моря в обеспечении промысла и воспроизводства кеты неодинаков (табл. 106, 107). Самой крупной по запасам кеты рекой на побережье является Охота. ТакLLLLLL LLLLL LL LLLLL LLLL LLLLLLL реках, как Пенжина и Уда, но фактических сведений по авиаучетам производителей кеты в них нет или они фрагментарны. Имеется члишь относительная оценкаролиэтих рек в воспроизводстве кеты Тугуро-Чумиканского района: р. Уда — 40—50%, p. Тугуp — 30—40% (Кульбачный, 2007). Эти два десятка рек (см. табл. 106, 107) составляют основу нерестового и промыслового запаса охотоморской кеты.
Горбуша. Ее воспроизводство в реках материкового побережья Охотского моря осуществляется в основном за счет естественного нереста природных популяций. На рыбоводных заводах ее доля в воспроизводстве составляет около 18% (Хованская, 2008). На материковом побережье Охотского моря доминируетлетняя форма горбуши. Ходна нерестпроисходит в конце июня—июле, основной нерест — в августе. Однако в некоторых реках (Гижига, Ола, Тауй) имеются группировки летней горбуши раннего и позднего хода на нерест.
В бас. p. Гижига установлено наличие двух экологических форм горбуши-ранней, совершающей нерестовую миграцию в течение июня, и поздней, заходящей на нерест в июле. Эти экологические группировки статистически достоверно различаются по биологическим показателям, темпуроста, структуречешуи (Марченко, 2001). Наличие ранней и поздней группировоклетней горбуши установлено и для других рек североохотоморского побережья — Олы и Тауя (Марченко, 1999).
Воспроизводство горбуши происходит в бассейнах более 90 основных рек, впадающих в Охотское море. Нерестится горбуша в центральной части мелководных русловых плесов и в концеплесов перед перекатами, где скорость руслового потока возрастает, при этом увеличивается инфильтрация воды валлювий нерестовых бугров, что повышает Проточность воды и выживаемостъ эмбрионов в нерестовых гнездах. Нерест горбуши отмечени в притоках первого и второго порядков. Предпочитаемая глубина 0,5-0,7 м, инoгда до 20 м. Инкубация икры горбуши происходит вводах подруслового потока, поэтому в конце сентября температура в нерестовых буграх снижается до 4—5°С, а в III декаде октября приближается к 0°C.
Как установлено И.С. Головановым (1982), величина нерестовых бугров североохотоморской горбуши меньше, чем в других районах Дальнего ВоcToka (Ky"3 HelloB kM. M., 1928; KaraHoBский, 1949; Воловик, 1967б; Рухлов, 1972; Чупахин, 1973; Смирнов А.И., 1975). Средняя длина бугра североохотоморской горбуши составляет 96, ширина — 68 см, площадь — 0,62 м". Исходя из этого определена оптимальная плотностъ заполнения нерестилищ-около 300 рыб на 100 м" (Голованов, 1982). По биологическим показателям охотоморская горбуша ближе всего стоит к горбуше Западной Камчатки, Kora CaxaJIMHa, AMypa (Ta6Jı. 108), To есть для нее характерны те же параметры, что и для других популяций горбуши, воспроизводящихся в бассейне северной части Охотского моря. Поздняя раса горбуши (Иванков, 1971) и горбуша южных областей азиатской части ареала крупнее (Курилы, Приморье). Наиболее крупным габитусом и
Таблица 10б. Удельный вес основных лососевых рек северного побережья Охотского моря в обеспечении промысла и естественного воспроизводст
107. Удельный вес основных лососевых рек побережья Охотского района в обеспечении промысла и естественного воспроизводства кеты
Среднемного- Доля Река летний FILOL= в воспроиЈход. тыс. рыб" | водстве, % Иня 87.7 6,0. Ульбея |013 3.24 KyxTyÄ 3504 122 Охота 22074 70,68 Урак 143, 458 Улья 109, 3.49 Все реки 31230 99.22
района
"Для рек побережья Охотского района взяты данные с 1986 по 2000 г.
плодовитостью отличается североамериканская горбуша Аляски, Британской Колумбии и южных штатов Вашингтон и Орегон (см. табл. 108).
На североохотоморском побережье основными горбушовыми реками являются Гижига, Наяхан, Вилига, Ола и Яна. Доминируют по запасам Гижига и Ола, доля которых в подходах и воспроизводстве составляет более 44% (табл. 109).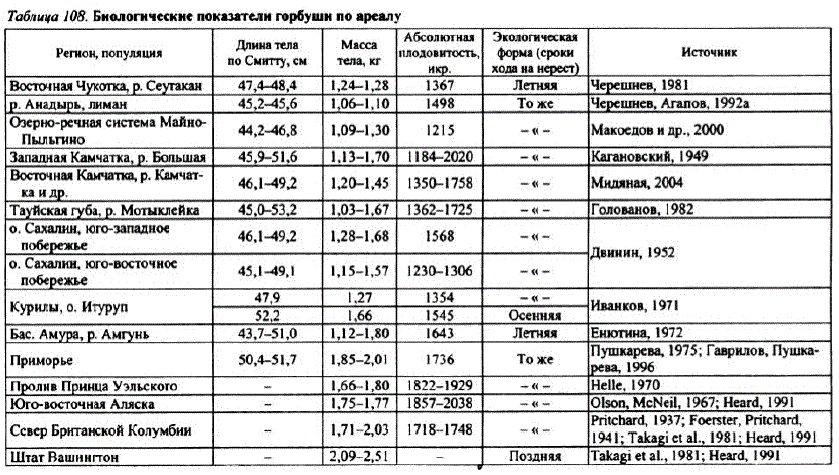
В Охотском районе запасы горбуши менее значимые, в среднем в 4,6 раза меньше, чем насевероохотоморском участке побережья. Основными реками по запасам горбуши здесь являются Иня, Кухтуй, Охота, Урак и Улья, их доля относительно общей величины подхода в среднем приближается к 90% (табл. 110).
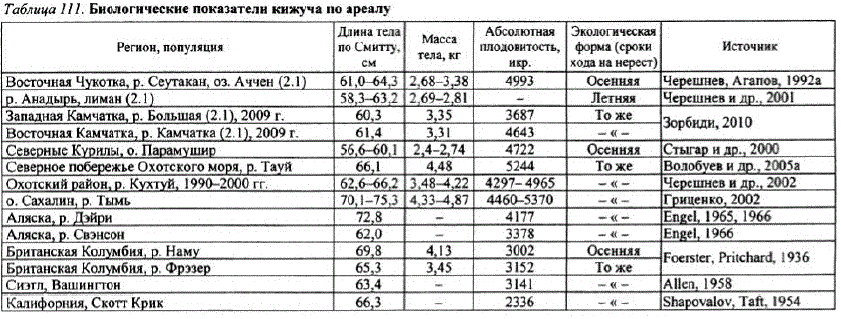
Кижуч. Его воспроизводство на материковом побережье Охотского моря поддерживается в основном за счет природных популяций. Его доля в заводском воспроизводственевелика и составила в среднем за все годы 4,4%. Более высокая численность кижуча отмечается в реках Охотского района: здесь его подходы в 1,5—2 раза больше, чем на североохотоморском побережье. На всем охотоморском побережье преобладает поздняя темпоральная форма кижуча, заходящая на нерест во второй половине августа—сентябре и размножающаяся в октябреноябре на выходах грунтовых вод, в ключах, затонах, лимнокренах.
Кижуч материкового побережья Охотского моря крупнее кижуча обоих побережий Камчатки и северных Курил. По размерноBCCOBEM IOKaaTCJAM H LOOBHтости он более сходен с сахалинским кижучем (табл. 111). Североамериканский кижуч при сходных размерах тела имеет более низкую плодовитость, что было отмечено ранее А.И. Смирновым (1975). У североамериканского кижуча наблюдается снижение плодовитости в направлении с севера на юг (Drucker, 1972), для азиатского кижуча такой закономерности не отмечено, На североохотоморском побережье подавляющее количество кижуча (более 98%) добывается и воспроизводится в четырех основных реках (табл. 112). Наибольшее значение имеет р. Тауй. В Охот
Таблица 109. Удельный вес основных лососевых рек северного побережья Охотского моря в обеспечении промысла
и естественного воспроизводства горбуши
ском районе основную роль в воспроизводстве и промысле кижуча играют три реки: Иня, Кухтуй и Охота. Их доля в оценке подходов кижуча составляет около
86% (табл. 113). Таким образом, на материковом побережье Охотского моря можно выделить две основные реки, за счет которых поддерживается воспроизводство и промысел кижуча, — Тауй и Охоту. Доля каждой из них в обеспечении подходов кижуча превышает 40%.
Нерка. Воспроизводится в некоторых реках побережья. Наибольшей численности она достигает в Охотском районе, где заходит на нерест в озерноречные системы таких рек, как Иня, Кухтуй, Охота. На северовосточном участке побережья она размножается в бас, pp. АвеKoBa, TM»KMTa, HasHxaH, OJIa, Tayñ и др. Нерестреофильной нерки отмечен в рр. Гижига, Быструха (Волобуев, Рогатных, 1984; Волобуев, Марченко, 2004). Макси-" мальной численности достигает в бас. р. Охота. В 1930-е гг. только в бас. оз. Б. Уегинское насчитывалось до 100 тыс. производителей (Никулин, 1975). При постройке нерестовых гнезд выбиpaeт yчacтки с выходами грунтовых вод как в озерах, так и в реках (Волобуев, Рогатных, 1997). В реках нерка размножается включевых протоках, в озерах строит гнезда на литорали в местах выходов грунтовых вод на глубине от 0,3—0,5 до 5 м. Из особенностей воспроизводства нерки отметим, что поJla BJ1 HH0LII a H HaCTb CC. B. B0/|0 CM a X материкового побережья Охотского моря размножается в озерах (более 97%). На Чукотке, напротив, доминируетреофильная форма нерки (более 95%) (Голубь, 2003).
Из табл. 114 видно, что лимнофильная нерка несколько крупнее ее речной формы, также нерка северо-восточной части материкового побережья несколько крупнее нерки, обита
ющей в юго-западной части побережья (р. Охота). Нерка из водоемов Магаданской области отличается и более высокой абсолютной плодовитостью. Особенно выделяется по этому показателю Среднемного- IloITH реофильная нерка из р. Гижига, Река ೫೫೦೫ || 880CIP೦ಗ್ಧ плодовитость которой приближа- ход, тыс. рыб" |водстве, % ется к плодовитости чукотской Яма 15.3 1949 нерки (см. табл. 114). OJIa 6,3 20,76 Регулярных авиаучетов нер- Яна 12.9 1643 ки в бассейнах рек Охотского рай- Тауй 3.7 40.38 она не проводилось, И.Ф. Прав- Все реки района 78.5 97.06
дин (1940) сообщало заходе на нерест только в одну Уегинскую озерно-речную систему в начале 1930-х гг. до 100 тыс. нерки. Имеются указания О.А. Никулина (1975) на то, что численность нерки в Уегинской озерно-речной системе в конце 1960-х гт колебалась в широком диапазоне — от 0,3 до 20 тыс. рыб. Более поздние Река ံးဖူးဖူ B စီးနင်္ဂါးnисследования (Пономарев, 2008) ход, тыс. рыб" | водстве, % показали, что запасы нерки в бас. ина .13 باشند р. Охота с 2003 по 2008 г. увели- VILőe 7 чились с 48 до 100 тыс. рыб, а в льоея mr. 궁중 бас. р. Иня на нерестилищах от Кухтуй 44,8 22.90 мечено 2—3 тыс. рыб этого вида. Охота 96.4 49.28 В бас.р. Олав конце 1990-х гг. (Пу- Урак 45 2,30 зиков, 1998) подходы нерки дости- Улья 52 2,66 гали 10 тыс. рыб, однако в послед- все реки района 1956 98.66
ние годы численность ее значительно сократилась. По результатам обследования в 2010 г. бас.
Таблица II.2. Удельный вес основных лососевых рек северного побережья Охотского моря в обеспечении промысла и естественного воспроизводства кижуча
"Для рек североохотоморского участка побережья взяты данные о подходах с 1996 по 2010 г.
Таблица 113. Удельный вес основных лососевых рек Охотского района в обеспечении промысла и естественного воспроизводства
"Для рек побережья Охотского района взяты Ila HHe c 1986 to 2000,
 оз. Киси, которое является основным нерестовым водоемом нерки в бас. p. Ола, численность ее производителей в конце августа определена всего внесколько сотен рыб. Достоверных сведений одинамике подходов и пропуска нерки в водоемы материкового побережья нет, поэтому трудно оценить их роль в воспроизводстве вида. Однако можно с уверенностью говорить о том, что pp. Охота и Кухтуй являются наиболее значимыми для воспроизводства нерки на материковом побережье Охотского моря.
оз. Киси, которое является основным нерестовым водоемом нерки в бас. p. Ола, численность ее производителей в конце августа определена всего внесколько сотен рыб. Достоверных сведений одинамике подходов и пропуска нерки в водоемы материкового побережья нет, поэтому трудно оценить их роль в воспроизводстве вида. Однако можно с уверенностью говорить о том, что pp. Охота и Кухтуй являются наиболее значимыми для воспроизводства нерки на материковом побережье Охотского моря.
Запасы охотоморских лососей по отношению к дальневосточным (2000—2011 гг.) невелики, их доля колеблется от 4,0 до 10,0% и в среднем составляет 6,9%. Однако следует отметить, что материковое побережье Охотского моря является крупным нерестовым комплексом тихоокеанских лососей на азиатской части нерестового ареала. Нерестовый фонд имеет хороший потенциали позволяет вместить гораздо больше лососей, чем их воспроизводится здесь сейчас.
Касаясь показателей биологической структуры, в целом можно отметить, что популяции охотоморских лососей имеют некоторые особенности. Так, например, кета представлена двумя сезонными формами и экотипами, тяготеющими к разным биотопам — ключевым и русловым.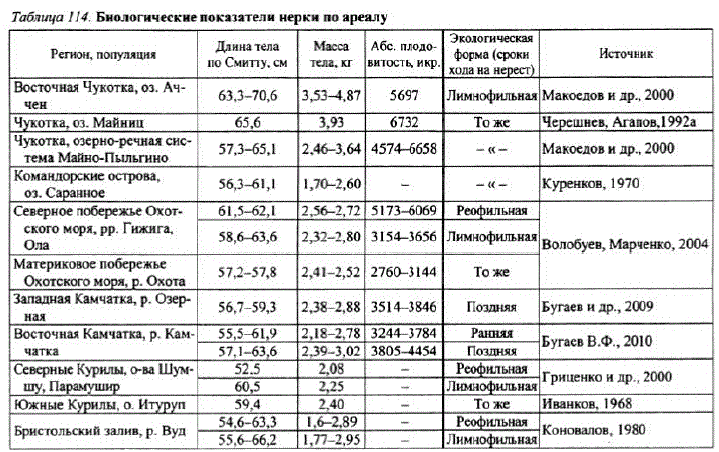
Горбуша отличается мелкими размерами именьшими, чем в других дальневосточных регионах, параметрами нерестовых бугров. KM»KyuH представлен
одной сезонной (осенней) формой и характеризуется более крупным габитусом, чем его представители из других регионов, уступая только сахалинскому.
Из двух форм нерки доминируетлимнофильная. Однако в целом биологические
Oncorhynchus.
характеристики всех видов лососей материкового побережья Охотского моря
леи рода
не выходят за пределы известных видоспецифичных параметров представите