НЕКОНТРОЛИРУЕМЫЙ ОТБОР В МАТОЧНЫХ СТАДАХ СЕМГИ (SALMO SALAR L.)
Множество популяций атлантического лосо-
ся (семги) поддерживается в настоящее время
путем искусственного разведения. В рыбовод-
ных хозяйствах многих стран создают маточные
стада рыб этого ценного вида, как с целью со-
хранения генофонда отдельных популяций, так
и для того, чтобы обеспечивать посадочным ма-
териалом товарные хозяйства. Однако при ре-
шении задачи создания маточных стад, предна-
значенных для сохранения природного гено-
фонда, ученые и рыбоводы сталкиваются с це-
лым рядом проблем.
Так, при искусственном воспроизводстве ат-
лантического лосося довольно часто происхо-
дило ненаправленное изменение частот некото-
рых генов (по сравнению с таковыми для диких
популяций), что было обусловлено недостаточ-
ным числом производителей, использованных в
рыбоводном процессе (Cross, King, 1983; Stahl,
1983; Verspoor, 1988; Crozier, Moffett, 1989;
Koljonen, 1989; Столь, 1991; Kazakov, Titov,
1993; Henricson et al., 1995; Tessier et al., 1995,
1997; Crozier, 1997; Danielsdottir et al., 1997;
Martinez et al., 2000; Koljonen et al., 2002; Saisa et
al., 2003). Однако в настоящее время разработа-
на система мер, позволяющих предотвратить
или, по крайней мере, существенно замедлить
дрейф генов (Cross et al., 1993; Дихнич, 2004).
Другая проблема, возникающая при искусст-
венном выращивании молоди атлантического
лосося – так называемый неконтролируемый
отбор (Никоноров и др., 1989), то есть отбор на
генетическую адаптацию к искусственным ус-
ловиям обитания (Казаков и др., 1987; Васин,
1988; Офицеров и др, 1989; Crozier 1998; Арта-
монова, Махров, 2002). О его действии во время
выращивания производителей в искусственных
условиях свидетельствует повышенная скорость
роста и созревания у потомков (Jonasson, 1993;
Kallio-Nyberg, Koljonen, 1997; H. King, pers.
comm., in: Thorpe, 2004), хотя на уровне отдель-
ных генов такой отбор показан не был.
И, наконец, не всегда удается избежать се-
лективности в процессе отлова производителей
для искусственного воспроизводства, поскольку
разные особи могут заметно различаться по
своим рыбоводным параметрам. Между тем, для
тихоокеанских лососей показано, что группы
рыб разного размера или срока хода в реку мо-
гут различаться также и по своим генетическим
характеристикам (Алтухов, 1974). Ряд авторов
полагает, что аналогичные явления могут наблю-
даться и у атлантического лосося (Дирин, 1977;
Riddell, 1986; Зелинский, Смирнов, 1987; Динами-
ка..., 2004), недавно получены прямые доказатель-
ства этого (Consuegra et al., 2005).
Целью настоящей работы было изучение не-
контролируемого отбора в ходе формирования и
выращивания маточных стад из потомков про-
изводителей реки Кереть (бассейн Белого моря)
на двух рыбоводных заводах Карелии.
Материалы и методы
Природная популяция. Река Кереть располо-
жена на западном побережье Белого моря
(рис. 1,А). Популяция атлантического лосося,
обитающая здесь, включает в себя производите-
лей как летней, так и осенней расы. До 1990 го-
да численность нерестового стада этой реки со-
ставляла несколько тысяч производителей. Од-
нако после того как в реку попал паразит
Gyrodactylus salaris (предположительно в 1990
году), популяция лосося резко сократилась, на
некоторых порогах дикая молодь встречается
единично (Шульман и др., 1998).
Заводское воспроизводство. С 1967 года ста-
до атлантического лосося реки Кереть частично
поддерживается путем искусственного воспро-
изводства молоди на Выгском и Кемском рыбо-
водных заводах (рис. 1,А). Молодь получают от
диких производителей, ежегодно отлавливае-
мых в нижней части реки. Одновременно для
рыбоводных работ используют более 100 про-
изводителей, однако все они являются рыбами
летней расы, поскольку из-за климатических
условий рыбоучетное заграждение может функ-
ционировать только в летние месяцы.
Рис. 1. A: Бассейн Белого моря; 1– река Кереть,
2 – Кемский рыбоводный завод, 3 – Выгский
рыбоводный завод. B: Бассейн реки Кереть; 1 –
рыбоучетное заграждение, 2 – порог ниже за-
граждения (Морской порог), 3, 4 – пороги вы-
ше заграждения
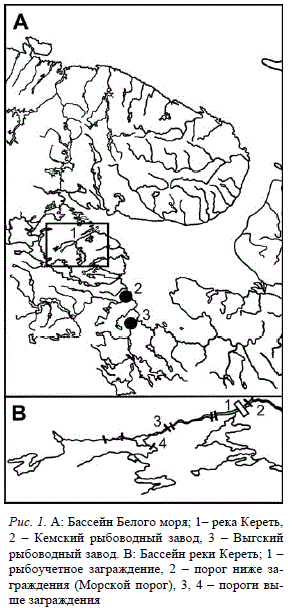
В первые годы рыбоводных работ в Кереть
иногда выпускали молодь, полученную от про-
изводителей из других рек (Артамонова, Мах-
ров, 2005), однако уже более трех десятилетий
чужеродную рыбу в реку не выпускают.
Маточные стада и их потомство. Для по-
лучения собственного маточного стада около
200 экземпляров молоди атлантического лосося
выращивали на Выгском рыбоводном заводе в
течение пяти лет. Все это время маточное стадо
содержали в пресной воде, несмотря на то, что
эти рыбы были потомками проходных особей
реки Кереть. Самки созрели на пятом году жиз-
ни, осенью 2000 года, и тогда же от рыб маточ-
ного стада было получено потомство (Крама-
ренко и др., 2002). Это потомство использовали
для восстановления полностью исчезнувших попу-
ляций атлантического лосося на территории Каре-
лии. Часть молоди (около 2/3) выпустили в реки
после первого года жизни, при этом на заводе была
оставлена только партия самой крупной рыбы.
На Кемском рыбоводном заводе в течение
трех лет выращивали экспериментальную пар-
тию атлантического лосося генерации 2000 года,
также полученную от производителей реки Ке-
реть. Более подробно этот эксперимент и его ре-
зультаты описаны нами в другой работе (Попова
и др., 2005).
Сбор проб. В работе исследованы выборки
молоди (включая потомство маточного стада) с
Выгского и Кемского рыбоводных заводов. Ино-
гда наряду с выборками живых рыб анализиро-
вали выборки погибших рыб (отход) из той же
группы особей (табл. 1). Кроме того, мы иссле-
довали диких пестряток, выловленных выше и
ниже рыбоучетного заграждения при помощи
электролова в 2001 и 2003-04 годах (рис. 1,В).
Образцы белых мышц и печени перевозили в
лабораторию в жидком азоте.
Электрофорез белков проводили по ранее
описанным методикам (Skaala et al., 1998). В ра-
боте исследовали пять локусов, кодирующих
белки, о которых было известно, что они поли-
морфны в других популяциях атлантического
лосося бассейна Белого моря (Skaala et al. 1998),
а именно: аспартатаминотрансферазу (AAT-4*),
эстеразу-Д (ESTD*), изоцитратдегидрогеназу
(IDHP-3*), сорбитолдегидрогеназу (IDDH-2*) и
малик-энзим MEP-2*).
Таблица 1. Частоты аллелей *100 в изученных выборках
Место сбора материала, Частоты аллелей * 100
генерация Возраст n ААТ-3* ESTD* IDDH-2* IDHP-3* MEP-2*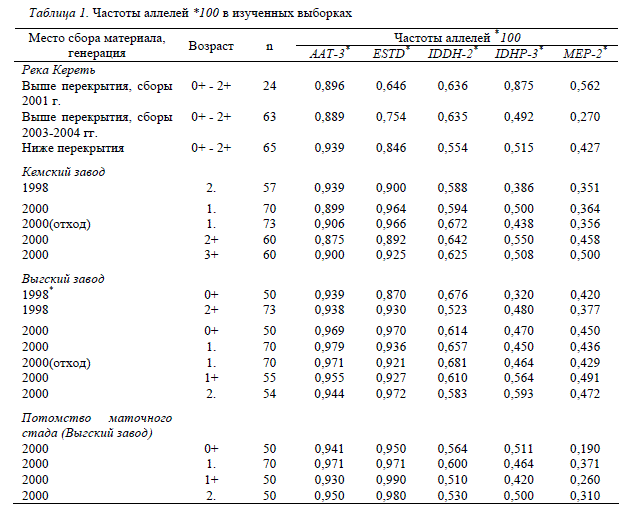
Примечание. * личное сообщение Е. К. Пономаревой.
Статистический анализ. Во всех выборках
оценивали частоты аллелей и соответствие рас-
пределения частот генотипов уравнению Харди-
Вайнберга. Для расчетов применяли компьютер-
ную программу BIOSYS (Swofford and Selander
1981). Гетерогенность частот аллелей оценивали
согласно (Roff, Bentzen, 1989) с помощью компью-
терной программы CHIRXC (Zaykin, Pudovkin,
1993).
Результаты
Генетическое разнообразие дикой молоди
реки Кереть. Все исследованные нами локусы
оказались полиморфными во всех изученных
выборках. Помимо аллелей, электрофоретиче-
скую подвижность которых принимали за
«100», мы выявили следующие аллельные вари-
анты: AAT-4*75, ESTD*94, IDHP-3*117,
MEP-2*135, IDDH-2*-90 и IDDH-2*40.
В таблице 1 представлены частоты аллелей
для двух групп диких рыб, – пойманных в
р. Кереть выше и ниже перекрытия. Судя по
распределению генотипов пяти исследованных
локусов, вся молодь, пойманная в отдельные
годы на некоторых верхних порогах, была пред-
ставлена потомками единственной пары произ-
водителей. При анализе полученных результа-
тов выборки, собранные на всех верхних поро-
гах в один и тот же сезон, объединяли.
Сравнение генетических характеристик
группы рыб, собранных в 2003-04 годах в верх-
нем течении с аналогичными характеристиками
группы молоди, собранной в это же время в
нижнем течении Керети, показало, что значи-
мых различий в частотах аллелей исследован-
ных локусов между ними нет. В то же время
совокупные выборки, собранные в разные годы
в верхнем течении р. Кереть, различались меж-
ду собой по частотам аллелей локусов IDHP-
3* (p < 0,001) и MEP-2* (p <0,001). Для ниж-
него течения Керети межгодовых сравнений
провести не удалось, поскольку выборка, со-
бранная здесь в 2001 году, была крайне мала
(включала всего 6 рыб).
Генетические различия между выборками
дикой и заводской семги. Частоты аллелей
*100 аллозимных локусов для выборок заво-
дской молоди семги представлены в таблице 1.
Чтобы выявить первоначальные генетические
различия между дикими и заводскими рыбами,
мы сравнили частоты в выборках сеголеток
(1998 и 2000 годов генерации) Выгского завода
и в выборке годовиков Кемского завода с часто-
тами аллелей в каждой из трех выборок диких
пестряток реки Кереть. Результаты анализа при-
ведены в таблице 2.
Временная динамика генофонда внутри
групп заводской молоди. Для того чтобы прове-
рить, имеют ли место генетические изменения
внутри каждой конкретной группы рыб в про-
цессе их выращивания в искусственных услови-
ях, мы ежегодно собирали выборки молоди из
каждой исследуемой группы, и сравнили эти
выборки между собой. При этом мы обнаружи-
ли значимые различия в частотах аллелей локу-
сов IDHP-3* (p<0,03) и IDDH-2* (p<0,026) меж-
ду двумя выборкам молоди генерации 1998 года
с Выгского завода. Одна из этих выборок была
собрана в 1999 году (возраст 0+), а другая – в
2001 году (возраст 2+). Значимых различий в
аллельных частотах между выборками живых
рыб и отхода ни в одной из исследованных
групп молоди нам обнаружить не удалось, од-
нако следует отметить, что для рыб генерации
1998 года анализ отхода мы не проводили.
Различия между генерациями. Значимые раз-
личия по частотам аллелей локуса IDHP-3* бы-
ли зарегистрированы между выборками рыб,
принадлежащих к генерациям 1998 и 2000 годов
как на Выгском (p<0,047), так и на Кемском
(p<0,03) заводах.
Таблица 2. Локусы, по которым отмечены значимые различия между выборками
молоди семги из р. Кереть и выборками, собранными на рыбоводных заводах
В первом случае мы сравнивали между собой
сеголеток, то есть молодь, которая практически
не подвергалась еще действию неконтролируе-
мого отбора, а во втором случае – двухгодови-
ков и трехлеток, что было обусловлено отсутст-
вием данных для рыб младших возрастов в слу-
чае генерации 1998 года. Кроме того, две выбор-
ки молоди, выращенной на Выгском заводе, раз-
личались также по частотам аллелей локуса
ESTD* (p<0,023). При этом значимых различий
между группами рыб одной генерации, но с раз-
ных заводов ни по одному из исследованных
локусов обнаружено не было.
Генетические особенности маточного стада
и его потомства. Мы обнаружили, что частоты
аллелей для локуса MEP-2* значимо различают-
ся в выборках сеголеток, полученных от произ-
водителей р. Кереть, отловленных на перекры-
тии, и сеголеток – потомков маточного стада
(p<0,001). При этом следует отметить, что обе
исследованные группы рыб принадлежали к ге-
нерации 2000 года и были выращены на Вы-
гском рыбоводном заводе. Сравнение между со-
бой всей совокупности выборок молоди разного
возраста, являющейся потомством маточного
стада, показало, что между выборками сущест-
вуют значимые различия в частотах аллелей ло-
куса MEP-2* (p<0,032).
Выборка рыб генерации 2000 года, возраста
3+, выращенных на Кемском заводе, значимо
(p<0,037) отличалась от выборки годовиков той
же генерации: частота «медленного» аллеля
MEP-2* здесь выросла (подробности см.: Попо-
ва и др., 2005).
Обсуждение
Состояние генофонда семги популяции
р. Кереть. В выборках, собранных на некото-
рых порогах, расположенных выше перекрытия,
распределение генотипов молоди позволяло
иногда предположить, что здесь нерестилась
только одна пара лососей. Таким образом, гене-
тический анализ позволил зарегистрировать не-
достаток производителей в верховьях реки, что
объясняется, по-видимому, тем, что в настоящее
время большая часть проходных рыб изымается
на перекрытии для заводского воспроизводства
или вылавливается браконьерами, а карликовые
самцы гибнут из-за заражения паразитом
Gyrodactylus salaris.
Вывод о недостатке производителей в вер-
ховьях реки косвенным образом подтверждает и
то обстоятельство, что выборки диких пестря-
ток, собранные в разные годы в верхнем течении
р. Кереть, различаются по частотам некоторых
генов. Кроме генетического дрейфа на это об-
стоятельство указывает также присутствие в ре-
ке гибридов семги и кумжи (Махров и др., 1998),
которые, помимо Керети, были обнаружены на-
ми и в соседней реке Нильма, где численность
популяции семги также сильно уменьшилась,
хотя и по другим причинам (Пономарева и др,
2002).
Исходно между дикой и заводской молодью
имелись значимые генетические различия. Наи-
более ярко они проявлялись для локуса ESTD*,
о чем свидетельствуют данные, представленные
в таблице 2, а также различия, обнаруженные
между выборками, собранными в реке и на Вы-
гском заводе в 1995 году (наши неопубл. дан-
ные). Скорее всего, эти различия объясняются
тем, что для заводского воспроизводства исполь-
зуют только рыб определенного срока хода. Та-
кое предположение представляется тем более
обоснованным, что ранее были отмечены разли-
чия по частотам аллозимов между группами
производителей, мигрирующих в другие реки
бассейна Белого моря в разное время. Различия
касались, в том числе, и локуса ESTD* (Семено-
ва, 1988).
Показательно, что в последние годы частоты
аллельных вариантов разных генов у дикой мо-
лоди Керети становятся более сходными с час-
тотами, характерными для заводской молоди.
Это связано, по-видимому, с тем, что естествен-
ное воспроизводство семги в Керети обеспечи-
вается в значительной степени рыбами заводско-
го происхождения.
Временная динамика генофонда внутри групп
заводской молоди. Мы исследовали две генера-
ции атлантического лосося (1998 и 2000 годов) с
Выгского рыбоводного завода, собрав выборки
рыб несколько раз за период выращивания (табл.
1). Для обеих генераций молоди было обнару-
жено, что частота «быстрого» аллеля IDHP-3*
уменьшается в выборках рыб старших возрастов,
причем в случае генерации 1998 года различия в
аллельных частотах между сеголетками и трех-
летками становились значимыми.
Интересно, кроме того, что сеголетки
1998 года генерации имели значимо более высо-
кую частоту «быстрого» аллеля IDHP-3* по
сравнению с молодью того же возраста, но
2000 года генерации. Исходя из того, что в на-
стоящее время заводское воспроизводство в по-
пуляции реки Кереть существенно преобладает
над естественным и составляет около 90%, нель-
зя исключить, что наблюдаемая тенденция явля-
ется следствием отбора против «быстрого» алле-
ля в заводских условиях, в результате которого
частоты аллелей этого локуса изменились уже и
у производителей.
С достаточно высокой частотой «быстрый»
аллель IDHP-3* встречается только в популяци-
ях, обитающих вдоль западного берега Белого
моря (Семенова, 1988; Офицеров и др, 1989;
Kazakov, Titov, 1993; Skaala et al., 1998) и весьма
вероятно, что его присутствие здесь является
следствием локальных адаптаций к условиям
некоторых рек. В заводских условиях присутст-
вие данного аллеля снижает, по-видимому, жиз-
неспособность его носителей, поскольку умень-
шение его частоты в выборках рыб старших воз-
растов регистрировали и ранее, а именно, – у
молоди семги реки Лувеньга, выращиваемой на
Кандалакшском рыбоводном заводе (Офицеров
и др., 1989).
Следует принять во внимание, что отбор по
другим аллозимным локусам, возможно, также
имеет место на рыбоводных заводах, однако мы
не можем выявить его из-за небольшого объема
исследуемых выборок. Так, в предыдущем ис-
следовании мы выявили отбор против гетерози-
гот по локусу ESTD* на Умбском рыбоводном
заводе в летний период, когда наблюдалась мас-
совая гибель рыб, сопутствовавшая аномально-
му повышению температуры воды (Артамонова и
Махров, 2002). Ранее другими исследователями
было отмечено изменение частот аллелей локуса
AAT-4* в ходе выращивания молоди на Нарвском
рыбоводном заводе в Ленинградской области (Ка-
заков и др., 1987).
Тем не менее, к сообщениям, касающимся
отбора против конкретных аллелей в заводских
условиях, следует подходить с осторожностью.
Наша практика показывает, что при работе с
заводской молодью большое значение имеет
методика сбора материала. Группы рыб, содер-
жащиеся в разных бассейнах, могут оказаться
потомками двух разных групп производителей,
состоящих не более чем из десятка особей каж-
дая, и при этом можно наблюдать порой значи-
мые различия в частотах аллельных вариантов
между молодью из разных бассейнов. Ошибок в
оценке частот удается избежать только при сбо-
ре материала таким способом, который гаранти-
рует равномерную представленность потомства
всех имевшихся производителей.
В связи с этими обстоятельствами затрудни-
телен также и анализ литературных источников
с целью выявления отбора на рыбоводных заво-
дах: анализируемые данные должны давать воз-
можность отсекать влияние дрейфа генов, гаран-
тировать отсутствие селективности при сборе
материала и его «загрязнение» рыбами иного
происхождения.
По этим причинам представляется сомни-
тельным, например, предположение об отборе в
заводских условиях против одного из аллелей
локуса триозофосфатизомеразы TPI-3* (оши-
бочно названного в работе TFI-3*) (Новиков,
Кузьмичев, 2003). Здесь при анализе литератур-
ных данных авторы ошибочно относят к заво-
дским те выборки, которые охарактеризованы в
оригинальной работе (Bourke et al., 1997) как
выборка диких рыб и выборки смешанного про-
исхождения.
Критически следует отнестись и к сообще-
нию о снижении уровня гетерозиготности по
ряду аллозимных локусов у молоди семги в ус-
ловиях Кандалакшского рыбоводного завода
(Евсюков и др., 2002), поскольку при анализе
авторы объединяли выборки рыб разных генера-
ций. Различия в частотах между выборками мо-
лоди, происходящей от разных производителей,
при этом не учитывались, а потому, в данном
случае, наблюдавшийся недостаток гетерозигот
объясняется, скорее всего, не отбором против
таких особей, а эффектом Валунда.
Генетические особенности потомков заво-
дских производителей и рыб из формируемых
маточных стад. Отличительной особенностью
потомства маточного стада, выращенного на
Выгском заводе, является аномально высокая
частота «быстрого» аллеля локуса MEP-2*. Ряд
фактов свидетельствует об адаптивном значении
полиморфизма по этому локусу. В частности,
было показано, что носители разных генотипов
MEP-2* отличаются друг от друга по скорости
роста и развития (обзор: Verspoor et al., 2005),
хотя различий по выживаемости между ними
ранее известно не было. Нами такие факты были
выявлены.
В процессе роста рыб частота «быстрого» ал-
леля во всех исследованных группах заводской
молоди имела тенденцию к уменьшению за
единственным исключением: она значимо воз-
росла у потомства маточного стада, когда после
сортировки молоди для доращивания на заводе
была оставлена только 1/3 часть рыб, причем
самых крупных.
Таким образом, имеются все основания пред-
полагать, что носители «быстрого» аллеля
MEP-2* имели в заводских условиях понижен-
ную жизнеспособность, однако именно они от-
личались высокой скоростью роста и были ото-
браны в свое время при создании маточного ста-
да. Ранее высокие частоты «быстрого» аллеля
MEP-2* были отмечены у линий лососей, пред-
назначенных для товарного выращивания
(Youngson et al., 1991) и прошедших отбор на
высокую скорость роста (Cross at al., 1993).
Избирательная гибель носителей «быстрого»
аллеля в условиях Кемского и Выгского заводов
объясняется, возможно, тем, что оба завода рас-
положены значительно южнее Керети: в литера-
туре имеются данные о том, что в реках с более
теплой водой преобладает «медленный» аллель
(обзор: Verspoor et al., 2005).
Что же касается преобладания «быстрого»
аллеля MEP-2* в потомстве маточного стада, то
помимо сортировки рыб по размерам, опреде-
ленную роль могло сыграть и то обстоятельство,
что при создании маточного стада на Выгском
заводе потомков проходных производителей со-
держали до созревания в пресной воде.
Анализ как литературных, так и собственных
данных показывает, что в пресноводных попу-
ляциях атлантического лосося «быстрый» аллель
MEP-2* представлен более широко, чем в про-
ходных. Например, в жилой популяции лососей
реки Намсен (Норвегия) он встречается с более
высокой частотой, чем в проходной популяции
этой реки (Vuorinen, Berg, 1989). Высокая часто-
та данного аллеля выявлена в выборках молоди
лосося из притоков некоторых озер бассейна
Белого моря (Tonteri et al., 2005; наши неопубл.
данные). Таким образом, вполне вероятно, что
некоторые генетические процессы, имеющие
место в процессе создания пресноводного ма-
точного стада, могут служить моделью, описы-
вающей формирование жилых популяций атлан-
тического лосося.
Доказательство адаптивности генетическо-
го полиморфизма. Уже несколько десятилетий в
литературе продолжается дискуссия об адаптив-
ности или нейтральности генетического поли-
морфизма (монографии: Левонтин, 1978; Киму-
ра, 1985; Алтухов, 2003). Нами и нашими колле-
гами получены экспериментальные свидетельст-
ва того, что полиморфизм, выявленный у атлан-
тического лосося по всем пяти изученным локу-
сам, кодирующим белки, может иметь адаптив-
ное значение.
Проблема сохранения генофонда в процессе
разведения атлантического лосося с целью
сохранения популяций. С генетической точки
зрения сохранение популяции — это сохране-
ние набора адаптаций, типичных для нее. В
нашем исследовании мы представили доказа-
тельства того, что в процессе искусственного
разведения атлантического лосося с целью
поддержания популяции, в группе искусствен-
но выращиваемых рыб может происходить не-
контролируемый отбор. Эта проблема весьма
актуальна для российских рыбоводных заво-
дов. Из-за низкой температуры воды рыбовод-
ный цикл здесь оказывается продолжительным
(более двух лет) и за это время сдвиг аллель-
ных частот в группах заводской молоди иногда
достигает значимых величин.
Таким образом, если естественный нерест в
реке по каким-то причинам не эффективен и по-
пуляция поддерживается, в основном, за счет
заводского воспроизводства, с течением времени
возможно существенное изменение генофонда
популяции. При этом не исключено, что если
сдвиг превысит некоторое критическое значе-
ние, то после прекращения искусственного вос-
производства такая популяция не сможет само-
воспроизводиться естественным путем, даже
если для этого будут созданы необходимые ус-
ловия.
Из этого следует, что для сохранения генети-
ческого разнообразия (а значит и присущего по-
пуляции набор адаптаций) в ходе искусственно-
го разведения, необходимы специальные меры
(обзор: Алтухов, 2003). Комплекс этих мер мож-
но назвать сохраняющей селекцией. Такая се-
лекция для атлантического лосося должна вклю-
чать, в частности, меры по предотвращению от-
бора рыб по размерам в процессе формирования
маточных стад и контроль за состоянием гено-
фонда в процессе выращивания рыб, особенно в
тех случаях, когда наблюдается массовый отход.
Необходимо также уделять внимание разра-
ботке мер, способствующих неизбирательному
повышению жизнестойкости рыб на всех стади-
ях рыбоводного цикла. Так, например, сохране-
нию генофонда семги при заводском выращива-
нии способствует кратковременное воздействие
на личинок лазерного излучения (Попова и др.,
2005).
В. С. АРТАМОНОВА, А. А. МАХРОВ, О. Н. ХОЛОД
Смотрите также