Разработка нового метода биотехники воспроизводства балтийской популяции атлантического лосося
Популяции атлантического лосося в Северо-Западном регионе утратили промысловое значение и поддерживаются за счет искусственного заводского воспроизводства.
Большинство лососевых рыбоводных заводов (ЛРЗ) заготавливает зрелых производителей на нерестилищах в ущерб естественному воспроизводству. Поэтому промысловая нагрузка ЛРЗ на нерестилища и в целом речной промысел лосося в период нереста является основной причиной снижения численности его популяций [1]. Второй важной причиной является низкий возврат производителей заводского происхождения, всего до 2% от общего количества выпущенной годовалой молоди. Это свидетельствует о необходимости повышения ее жизнестойкости, разнокачественности и разработки новой современной биотехники заводского воспроизводства в целом. Отечественный опыт воспроизводства атлантического лосося и зарубежные проекты сохранения его популяций указывают на необходимость сочетания рыбоводных эффектов заводского воспроизводства с естественным [1,2].
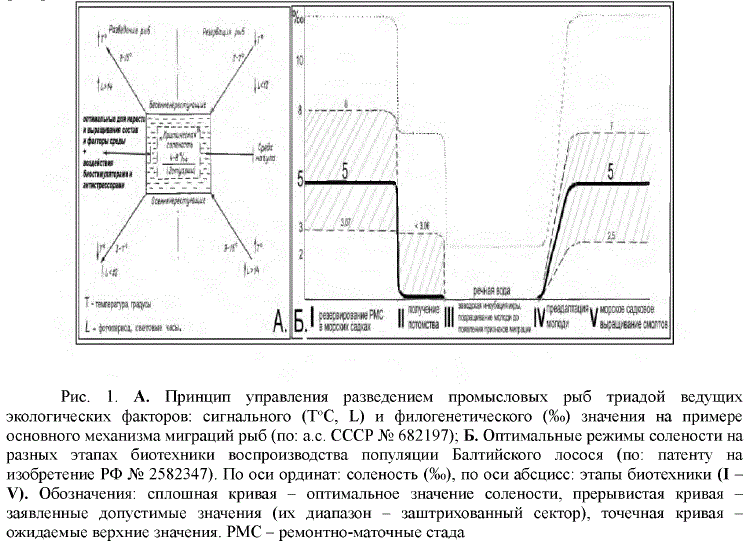
Поэтому с целью повышения эффективности заводского воспроизводства лосося разработан новый метод управления размножением, темпами роста, степенью развития и подготовленности (преадаптации) молоди к морскому образу жизни [3]. Он осуществляется путем массовой заготовки производителей в море, содержания маточных стад в морских садках, получения здесь потомства и, после заводского выращивания в реке личинок и молоди, доращивания заводских смолтов в солоноватой морской воде (рис. 1).
Материалы, методы и объекты исследования. Сравнительные результаты испытаний нового метода воспроизводства Балтийской популяции атлантического лосося приведены в таблице.
Мы считаем, что получение потомства от производителей в морской солоноватой воде на местах нагула и промысла имеет следующие основные преимущества: 1. Снятие промысловой зависимости с JIP3 и промысловой нагрузки с нерестилищ, что может исключить все виды речного браконьерства. 2. Объединение интересов всех видов воспроизводства и промысла с промышленным использованием производителей. 3. Снижение производственных потерь при содержании ремонтно-маточных стад (РМС) в оптимальной среде резервирования [1].
Доращивание молоди в солоноватой морской воде на местах нагула (пастбищах) с момента начала смолтификации также имеет ряд преимуществ:
1. Многократное усиление темпов роста при прочих равных условиях, особенно значительное с годовалого возраста. 2. Снижение производственных отходов в процессе смолтификации, приобретающем массовый синхронный характер, соответствующий природному, а также за счет исключения «речных» карликовых самцов. 3. Снижение основных производственных потерь в результате повышения выживаемости смолтов, адаптированных к среде нагула, и степени возврата «заводских» производителей.
Сокращение этапов биотехники непосредственно на JIP3 (рис. 1 Б) высвободит дополнительные производственные мощности для достижения необходимых масштабов и эффективности заводского воспроизводства. Новый метод впервые позволит сочетать искусственное воспроизводство с естественным в единый природно-промышленный комплекс воспроизводства [1, 3].
Преимущества вышерассмотренного первого этапа предложенного метода -формирование и эксплуатация РМС, получение потомства в солоноватоводной морской среде в природоохранном и рыбохозяйственном аспектах не нашли серьезных обоснованных противопоказаний в литературе и не вызвали возражений опрошеных нами специалистов. Очевидно, что на этом этапе особо необходимы постоянные мониторинг состояния производителей, предотвращение их выхода из садков и освежение состава РМС.
Однако выяснился ряд серьезных возражений против конечного этапа метода в виде доращивания заводской молоди в морской воде на местах нагула (пастбищах) с момента начала смолтификации:
1. Заводская молодь, сбежавшая из морских садков, нарушает экологическое состояние и генетическую структуру местных природных популяций, что отражено в решениях организации ФАО [4]. Поэтому Европейская комиссия по атлантическому лососю предложила прекратить компенсационные выпуски смолтов в Балтийское море, допуская только выпуски ранних стадий развития (до ранней молоди), при отсутствии естественного нереста [5]. Однако для увеличения промысловых уловов путем пастбищного рыбоводства допускается выпуск смолтов или пост-смолтов (Sea ranched salmon) в море.
2. Возможно нарушение хоминга у годовалых заводских смолтов после доращивания в морских садках. Однако мы считаем, что перевод конечного заводского цикла биотехники в море для усиления роста и выживаемости молоди вполне возможен, поскольку хоминг лососей генетически не закреплен, а импринтинг формируется уже в первое лето заводского выращивания личинок и ранней молоди с момента перехода на активное питание. Это доказано для тихоокеанских лососей [6], в частности опытами на сеголетках кижуча, выращенных в течение 1 месяца в бассейнах с добавлением 1М-гидроксиэтил-морфолина, либо в другом варианте опыта - фенетилового спирта [7]. После выпуска и 18-месячного нагула в море у подопытной молоди был получен яркий эффект управляемого («облигатного») хоминга - возвращение в реки с содержанием этих растворенных химикатов (соответственно: 95 и 92% возврата в обработанные каждым препаратом «чужие» реки).
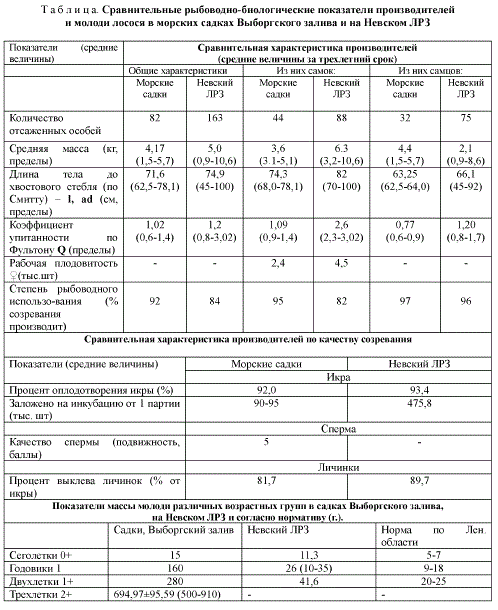
К настоящему времени установлено, что процессы импринтинга и хоминга осуществляются в скоплениях люлиберинергических (гонадолиберинергических) нейросекреторных клеток (ЛГ-РГ-НСК), которые в целом локализуются вблизи зрительных и обонятельных центров переднего и промежуточного мозга, преимущественно в 3-х отделах (рис. 2): 1) в ганглии терминального нерва (вблизи Organum vasculosum laminae terminalis -OVLT), между обонятельными луковицами и передним мозгом (Nucleus olfactorius - NOR), 2) в преоптической области, по сути в нейросекреторном преоптическом ядре гипоталамуса -ПЯ, (N. anterior periventricularis - NAP), 3) в передней части латерального ядра серого бугра гипоталамуса у многих видов костистых рыб (Nucleus lateralis tuberis - NLT, anterior).
У мигрирующих на нерест моноцикличных лососей (кеты и нерки) в разных отделах мозга установлена различная динамика синтетической активности ЛГ-РГ, а именно: усиление синтеза в области обонятельного нерва в NOR при заходе рыбы в низовья рек и смещение пика активности синтеза в преоптическую область (NAP) в период нереста на нерестилищах [6].
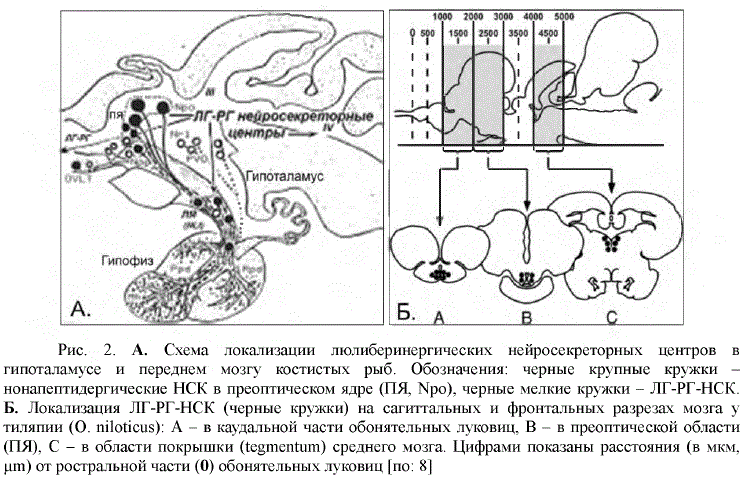
Установлено дифференцированное участие разных форм ЛГ-РГ и соответственно ЛГ-РГ-НСК в миграциях и нересте (соответственно биологической значимости хемо- и фоторецепции): NOR - в процессах импринтинга и хоминга и NAP (ПЯ) - на различных этапах полового созревания и нереста. При этом все формы ЛГ-РГ вовлекаются при нересте в модуляцию сезонного репродуктивного поведения, особенно его социальных форм, например агрессии у самцов, что определяется отрицательной обратной связью с уровнем содержания андрогенов (тестостерона, 11кето-тестостерона).
Задача выяснения природы «миграционного импульса» была поставлена еще проф. Н.Л. Гербильским и разрабатывалась его школой эколого-гистофизиологического направления ихтиологических и рыбохозяйственных исследований [9, 10]. Ведущая роль в детерминации миграционного поведения исходно отводилась гипоталамо-гипофизарной нейросекреторной системе (ГГНС; рис. 2А), выполняющей специализированные (водно-солевой обмен, тонус гладкой мускулатуры, нерестовое поведение) и генерализованные функции (метаболический гомеостаз организма). Важно, что активация ГГНС выражена в двух альтернативных формах: 1 - накопление в заднем нейрогипофизе (ЗНГ) нейросекреторных продуктов (аккумуляция их в ЗНГ) с активацией их синтеза и транспорта в преоптическом ядре, т.е. опустошения ПЯ от них - состояние «мобилизации» ГГНС (как латентное) и 2 - выведение нейрогормональных продуктов из ЗНГ в общий кровоток, т.е. опустошение ЗНГ от нейросекреторного материала (НСМ) - состояние активации ГГНС на уровне организма.
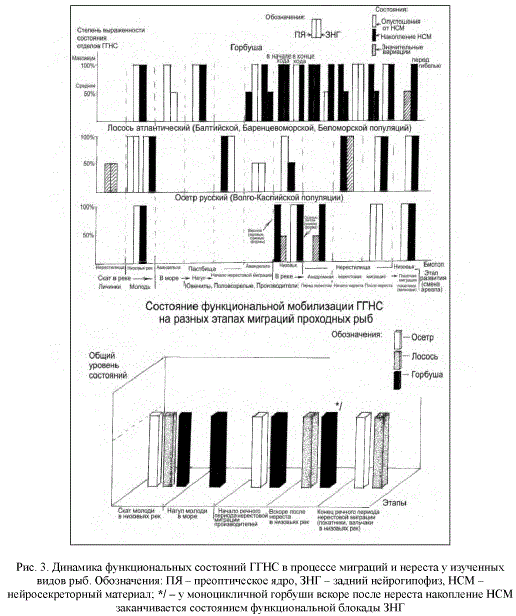
Эколого-гистофизиологический анализ участия ГГНС в осуществлении нерестовых миграций и нереста позволил установить, что ее нонапептидные нейрогормоны (аргинин-8-вазотоцин и изотоцин) инициируют нерестовое поведение и соматические перестройки, затем способствуют овуляции и спермиации, стимулируя сокращения гладких мышц гонад [1]. К завершению нереста оба нейрогормона, особенно вазотоцин, наиболее активно участвуют в защитно-приспособительных реакциях организма, направленных на преодоление состояния физиологического стресса, возникающего при нересте рыб, особенно на поддержание метаболического равновесия организма. Состояние физиологического стресса мы рассматриваем как конечное звено в последовательных этапных процессах, обеспечивающих явление прогрессирующего снижения степени эврибионтности проходных рыб в процессе полового созревания, миграции и нереста [1, 9, 10]. Прежде всего нонапептидные нейрогормоны ГГНС в комплексе с половыми и кортикоидными стероидными гормонами играют важную роль в детерминации нерестового миграционного поведения, создавая в ЦНС половую доминанту. Анализ собственных материалов с применением современных морфометрических методов оценки функционального состояния ГГНС у основных изученных ценных видов проходных рыб в процессе миграций и нереста (и сопоставление их с данными литературы) позволяет впервые представить следующую динамику ее активности на важнейших этапах их онтогенеза (рис. 3).
Из представленной гистограммы очевидно, что детерминирующим звеном, общим для различных форм миграций разных видов ценных промысловых рыб является состояние функциональной мобилизации ГГНС. Оно в принципе сходно с уже известными состояниями нижних звеньев гипоталамо-гипофизарно-висцеральных осей нейро-эндокринных взаимоотношений и, по-видимому, является общим и для всего нейроэндокринного комплекса. Их общая активация наступает уже после перехода в новую среду обитания и при нересте как результат стресса, необратимого (дисстресса) у моноцикличных рыб.
Наиболее перспективным направлением дальнейшего развития НИР представляется анализ механизмов усиления роста и выживаемости молоди в солоноватой среде путем изучения развития функциональной оси соматолиберин - соматотропин - соматомедин (и в его комплексе с ГГНС) современными морфометрическими методами и взаимодействия люлиберинергических и нонапептидергических центров (ЛГ-РГ НСК с НП-НСК), с конечной целью управления процессами импринтинга и хоминга (рис. 1 А, 2А).
Выводы. Для дальнейшей разработки и внедрения нового метода современной эффективной биотехники воспроизводства лосося непосредственно на ЛРЗ, и учитывая указанные возражения по морскому садковому выращиванию крупных смолтов, предлагаются к использованию системы замкнутого водоснабжения (УЗВ) рыбоводных заводов и рыбоводных хозяйств, впервые основанные на принципе внесезонного подземного гидрокондиционирования управляемой среды выращивания (авторское свидетельство СССР № 982614, патент РФ № 2400975). Поэтому первым этапом НИОКР является разработка оптимальной (модифицированной) среды для эффективного выращивания в УЗВ заводской молоди лосося, аналогичной уже разработанной нами для резервирования производителей промысловых рыб, осетровых и костистых (авт. свид. СССР № 965409).
Доктор биол. наук П.Е. ГАРЛОВ (СПбГАУ Этот адрес электронной почты защищён от спам-ботов. У вас должен быть включен JavaScript для просмотра.) Канд. биол. наук С.У. ТЕМИРОВА (СПбГАУ Этот адрес электронной почты защищён от спам-ботов. У вас должен быть включен JavaScript для просмотра.) Соискатель B.C. БУГРИМОВ (Сев.-Зап. Терр. Управл. ФАР, Этот адрес электронной почты защищён от спам-ботов. У вас должен быть включен JavaScript для просмотра.)