Состояние запасов и промысел кеты
Во второй половине XX в. численность кеты материкового побережья Охотского моря колебалась в широких пределах — от нескольких десятков тысяч в годы депрессии (конец 1960-х — начало 1970-х гг.) до 9 млн рыб в 2000-х гг. Основными факторами, определяющими запасы кеты, являются: эффективность
воспроизводства, сопряженная с воздействием на выживаемость потомства климатических условий как в пресных водах, таки в прибрежье; условия нагула в океане, определяющие уровень смертности (до 60—80%) (Старовойтов, 2003); влияние факультативных хищников пресных водиморского прибрежья, пресс берегового и океанического промысла, браконьерский вылов, уровень пропуска производителей на нерестилища.
Состояние запасов лососей материкового побережья Охотского моря анализировалось ранее (Костарев, 1964a, 1970в, Клоков, 1970, 1973а). Однако считаем целесообразным вернуться к рассмотрению этого вопроса, так как состояние стад лососей региона претерпело определенные изменения за прошедшие годы.
Представление о запасах охотоморской кеты могут дать сведения повылову. Данные о выловекеты на материковом побережье Охотского моря имеются за 100 лет: с 1908 по 2010 г. (Уловы., 1989; данные ФГУ «Охотскрыбвод»; Охотского территориального управления Росрыболовства). Естественно, сведения о вылове не могут служить мерилом, адекватным оценке запасов, так как коэффициент изъятия — величина непостоянная, тем не менее объемы вылова дают общее представление об уровне подходов лососей. В 1950—1960-е гг. уровень промыслового изъятия охотоморской кеты колебался от 32 до 77%, приближаясь в среднем к 60% (Костарев, 1970в).
В динамике численности кеты материкового побережья Охотского моря можно выделить несколько периодов: первый — время высокой численности, второй — снижение запаса и депрессия численности, третий — рости некоторое увеличение запаса, четвертыи — максимум численности за последепрессионный период. Затем наблюдался очередной спад уровня запасов, обусловленный общим снижением численности и мощным браконьерским прессом в 1990-е гг. Современный период состояния запасов характеризуется медленным ростом HPCCHHOCTBTECOB.
Первый период охватывает 30—50-е гг. XX в. — время благополучия стад охотоморской кеты, принимаемое нами за период оптимума запасов, когда численность ее подходов к побережью достигала 15—18 млн экз., а вылов в северовосточном районе (Магаданском) побережья колебался от 2 до 9 тыс. т, в югозападном (Охотском) — от 8 до 32 тыс. т. В дальнейшем чрезмерная эксплуатация кеты, совпавшая с рядом лет, неблагоприятных по условиям воспроизводства, привела к резкому снижению ее запасов. Для сравнения можно отметить, что в 1950-е гг. в Охотском промысловом районе в лососевую путинуработало до 160 неводов, а вылов кеты достигал 32 тыс. т (Historical..., 1979), тогда как к началу 1990-х гг. количество орудийлова составляло около 15, а вылов снизился до 2,1—3,6 тыс. т (табл. 39).
Сактивизацией в 1950-х гг. иностранного океанического промысла азиатских лососейуловы Японии, чьи суда вели их основной дрифтерныйлов в северозападной части Тихого океана, стали возрастать. К 1955 г. они сравнялись с российскими береговыми, а с 1957 г. стали значительно превышать их. Максимальный вылов Японией лососей азиатского происхождения был отмечен в 1958 г., когда он достиг 199 тыс. т, из которых более 70 тыс. т пришлось на долю кеты (Historical..., 1979). Максимальный зарегистрированный выловкеты Японией составил 99,5 тыс. т (1975 г.).
Следует отметить, что фактическая величина вылова лососей была на 30% и более выше данных официальной статистики вследствие косвенных потерь сырца при океаническом промысле (выпадение из сетей, выедание хищниками, травмированность рыб и др.) (Синельников, 1983).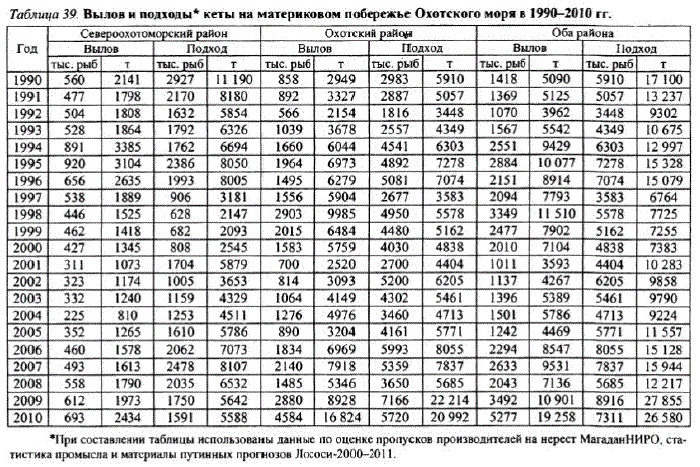
Интенсификация японского океаническоголова азиатских лососей сопровождалась снижением уровня отечественного прибрежного промысла. В результате мощного пресса иностранного морского промысла в сочетании с отечественным прибрежным и устойчивым дефицитом производителей на нерестилищах запасы кеты материкового побережья Охотского моря сократились почти на порядок (Костарев, 1983). Кроме того, снижение подходов лососей совпало с чередой лет, неблагоприятных для воспроизводства кеты в климатическом отношении в конце 1950-х — первой половине 1960-х гг. (суровые малоснежные зимы, обсыхание и промерзание нерестилищ). Это обусловило появление ряда неурожайных поколений в 1960-х гг. (Костарев, 1964б).
Чрезмерная эксплуатация запасов лососей азиатского происхождения привела в конечном итоге к подрыву ресурсов кеты, что выразилось в депрессии численности вида, начавшейся в конце 1960-х и продолжавшейся до середины 1970-х гг. Запасы охотоморской кеты в эти годы не превышали 10—15% от уровня лет оптимума. Депрессивное состояние запасов кеты на северо-восточном участке побережья отмечено с 1966 по 1976 г., в юго-западном районе (Охотском) — с 1968 по 1977 г. (рис. 33). Уловы кеты на северном побережье снизились до 40 т (1969, 1973 г.), в Охотском районе — до 110—160 т (1968, 1973 г.). Крайне нерациональный морской промысел лососей, когда потери биомассы за счет вылова неполовозрелых особей составляли до 30% (Костарев, 1978, Вронский, 1980), послужил основной причиной не только резкого сокращения общего запаса, но и значительного изменения внутривидовой структуры стадкеты. Любое нарушение структуры популяционных систем вследствие выпадения одного из звеньев или снижения генного разнообразия популяции (стада) может привести к нежелательным трансформациям, снижающим гомеостатический уровень и общую численность вида на участке ареала. Так, например, в реках Охотского района были подорваны запасы ранней кеты, на которой раньше (в 1940—1950-е гг.) базировался промысел. В 1970—1980-е гг. запасы кеты материкового побережья поддерживались в основном благодаря поздней форме вида (Волобуев, Тюрнин, 1995). Выживаемость поздней кеты выше, чем ранней, однако численность ее лимитируется наличием мест, пригодных для воспроизводства, — ключевых проток, лимнокренов, затонов с выходами грунтовых вод (Волобуев и др., 1990). Поэтому ее численность на материковом побережье Охотского моря не достигала высокого уровня. Ресурсы же ранней кеты, которая прежде составляла основу подходов и вылова, в эти годы были невысоки — около 6% от уровня подходов (Евзеров, 1983; Волобуев, Голованов, 2001).
Снижение запасовкеты отразилось и на урожайности ее генераций. В изменении численности отдельных поколений охотоморской кеты можно выделить те же периоды, что и в изменении общего запаса. Каждый из этих временных интервалов характеризуется разным уровнем выживаемостикеты. В периоддодепрессии (1956— 1963 гг.) имела место довольно высокая численность поколений (2,4—11,1, в среднем 5,4 млн рыб). Период спада запасов (1964—1973 гг.) — время резкого, в 2—3 раза, снижения численности поколений, когда их средняя величина составила 1,6 (0,6—4,2) млн рыб. В постдепрессионный период (1974—1987 гг.) наблюдался рост численности поколений, в среднем онадостигла 3,6 (1,7—5,6) млн рыб.
Показателем выживаемости поколений кеты служит кратность естественного воспроизводства, определяемая как отношение численности потомства (поколения) к численности родительского стада (Леванидов, 1969).
Среднемноголетний показатель кратности воспроизводства, рассчитанный для отдельных районов, составил: 1,81 для популяций кеты Гижигинской губы, 1,59 для рек Ямской губы и 1,56 для Тауйской губы. В среднем для северовосточного побережья кратность воспроизводства 27 поколений природных популяций ксты составила 1,61 при колебаниях от 0,31 до 4,4; для 32 поколений кеты Охотского района — 1,65 с размахом колебаний от 0,16 до 5,75. Показатель кратности воспроизводства имеет слабую обратную связь с численностью P??????? ? северо-востока побережья r = -0,47, p = 0,98; для юго-запада r = -0.13; p < 0,9). (а) и в Охотском промысловом районе (б)")
Рис. 33. Динамика подходов кеты на северном участке побережья Охотского моря (Магаданский регион) (а) и в Охотском промысловом районе (б)
Наиболее урожайными на северо-востоке побережья были поколения 1961, 1962, 1977, 1982 и 1985 г., прослеживающиеся в каждом из стад (гижигинском, ямском, тауйском). В юго-западном районе (охотское стадо) к урожайным относятся IOKoIeHHA 1956, 1961, 1969, 1973, 1975, 1977 H 1981 I. MakcHMa I. HoecxoIcTBo Io числу урожайных лет наблюдается между кетой тауйского и охотского стад, что, видимо, свидетельствует осходной динамике природных процессов вэтих районах воспроизводства. Четкая 4-летняя цикличность урожайных поколений по нечетным годам прослеживается для кеты гижигинского и охотского стадс 1969 по 1985 г. В остальных районах зависимости возникновения урожайных поколений отурожайных не наблюдалось. Однако отметим, что количество урожайных поколений в 1970— 1980-е гг. возрослов 6—8 раз по сравнению с годами депрессии.
B целом для кеты материкового побережья Охотского моря характерно расширенное воспроизводство. Численностъ дочерних поколений лишь в 7 из27 случаев была ниже численности родителей для северо-восточного участка побережья и в 11 из 32 поколений для Охотского района.
Заметный подъем уровня запасов охотоморской кеты начался с 1980-х гг., KOTA была знач ительно снижена квотаморского вылова JOCOceji oce BEBEECния в 1978 г. Россией 200-мильной экономической зоны. К 1983 г. морской выловлососей был сокращен до 42 тыс. т, а впоследующие годы — до 15 тыс. т (Зиланов, Спивакова, 1992). Параллельно с сокращением морского промысла происходило замещение относительной доли кеты в морских уловах с 60% в 1970-е гг. до 24—30% в 1980-е гг. С 1992 г. океанический промысел азиатских лососей был прекращен, и с тех пор реализация квот, выделяемых иностранным государствам, осуществляется в рамках межправительственных соглашений в пределах экономической зоны России.
Подходы североохотоморской кеты достигли максимума за последепрессионный период в 1990 г. (2,9 млн рыб). В соответствии с этим, к середине 1990-х гг. ее вылов в Магаданской области превысил 3 тыс. т. Затем стали снижаться как подходы, так и доля участия Магаданского региона в воспроизводстве кеты материкового побережья. Наименьшего уровня подходы охотоморской кеты достигли в конце 1990-х гг. (1998—1999 гг. — 0,6 млн рыб), а ее доля в общем береговом выловедальневосточной кеты снизилась до 13% (табл. 40).
Таблица 40 Доля (%) кеты материкового побережья в общем береговом вылове дальневосточной кеты (1) и доля кеты в общем вылове лососей в море (2)
Вылов
Главными факторами, обусловившими уменьшение ее запасов, явились интенсивный морской промысел на путях преднерестовых миграций (согласно экспертной оценке, морской вылов кеты материкового побережья Охотского моря в 1990-е гг. достигал 40%) и чрезмерный пресс берегового промысла, значительную долю которого составлял нелегальный лов в реках и на нерестилищах. В 1990-х гг. суммарное изъятие официального и браконьерского вылова в некоторых реках достигало 80—90%. Такое снижение запасов было сопряжено с резким усилением берегового браконьерскоголова, обусловленного социальной неустроенностью населения в период перехода страны к рыночным отношениям впервой половине 1990-х гг. При этом широко практиковалась сортировка рыб по половому признаку с изъятием самок из уловов, в результате чего искажалось соотношение полов нанерестилищах, и это негативно влияло на уровень и эффективность естественного воспроизводства стад. Опустошенные нерестилища, естественно, не могли обеспечить высоких возвратов, поскольку запасы североохотоморской кеты определяются состоянием ее природных популяций. Пропуск производителей на нерест сократился в несколько раз.
Исторически на материковом побережье Охотского моря существуют два основных промысловых района: Североохотоморский (участок побережья к северо-востоку от п-ова Лисянского до п-ова Тайгонос включительно) и Охотский (участок побережья к юго-западу от п-ова Лисянского до Аяна включительно). На протяжении 1970—1980-х гг. доля кеты в воспроизводстве и промысле обоих районов была примерно равной. Доля популяций североохотоморской кеты составляла в среднем 50,8%, кеты Охотского района — 49,2% запаса.
В начале 1990-х гг. мягкие многоснежные зимы и череда теплых в гидрологическом отношении лет (1993—1997) способствовали хорошей выживаемости и более высоким подходам кеты в Охотский район. К 1994 г. ее подходы в Охотский район составляли уже более 70%, а к 1998—2000 гг. возросли до 86—88% запаса всего п (см. табл. 39), что в абсолютном выражении составляло до 5—5,2 млн рыб. Затем с 2001 г. численность подходов стала снижаться и вылов уменьшился с 100 (1998 г.) до 2,5—3,1 тыс. т в 2001—2002 гт. К 2006—2007 гт. вылов снова увеличился до 7—7,9 тыс.т. Сростом запасов в Североохотоморском промрайоне общий подходкеты достиг 8,9 млн рыб, а вылов — 10,9 тыс. т.
Доля кеты материкового побережья в общем береговом вылове дальневосточной кеты также изменялась, что было обусловлено разным уровнем ее запасов в отдельных лососевых регионах (см. табл. 40). Роль охотоморской кеты в уловах дальневосточной кеты возросла в 1994—1998 гг. Это, очевидно, было обусловленоростом подходов кеты в Охотский район и снижением значения в подходах и воспроизводстве кеты Восточной и Западной Камчатки. Подходы кеты на обоих побережьях Камчатки увеличились с 1999 г., что, по-видимому, было сопряжено со снижением объемов ее вылова в море и усилением контроля за дрифтерным промыслом в северо-западной части Тихого океана (Лососи-2005) и ростом запасов сахалинской и южнокурильской кеты за счет искусственного воспрозводства. В соответствии с возросшими подходами увеличился и выловкамчатской кеты — с 3—4 до 26 тыс. т, сахалинской и амурской — до 20 тыс. т. На фоне роста запасов камчатской, амурской и сахалино-курильской кеты произошло снижение удельного веса охотоморской кеты в ее общем запасе Дальнего Востока (см. табл. 40). В целом среднемноголетняя доля выловакеты материкового побережья в ее общем береговом вылове за это время составила 32,9%. В отношении доли выловакеты дрифтерными судами при проведении научно-исследовательских работ и промысла по межправительственным соглашениям с Японией в российской экономической зоне можно сказать, что она является доминирующим видом, составляя в среднем 52% от общего вылова всех видов лососей.
При рассмотрении рис. 33 можно заметить, что до середины 1990-х гг. прослеживалась HETKasi TCHICH II HA роста BLOCOB KOTE KERK TO северному побережью, так и по Охотскому району. Рост запасакеты на северном побережье происходил более быстрыми темпами по сравнению с кетой Охотского района. К началу 1990-х гг. запасы охотоморской кеты увеличились в 2,4 раза по отношению к последепрессионному десятилетию. Современный уровень возвратов кеты в реки Охотского района составляет лишь 50—60%, в реки северного побережья — 50-70% от подходов лет оптимума численности. Абсолютная численность подходов выше в Охотском районе. На северном побережье наиболее высокая численность кеты отмечена в районе Гижигинской губы, куда в 1960-е гг. на нерест подходило до 2 млн рыб, второй позначимости — район Ямской губы (до 1,4 млн рыб), третий — Тауйской губы (до 0,6 млн рыб).
Несмотря на некоторое увеличение численности нерестовых подходов кеты, запасы ее еще далеки от оптимальных, поэтому в целях сохранения тенденции роста численности возвратов предусмотрен щадящий режим ее использования. В Охотском районе и реках зал. Шелихова промысловое изъятие кеты рекомендовано в зависимости от урожайности возвращающихся на нерест поколений с учетом обязательного пропуска на нерест оптимума производителей. Для североохотоморского участка побережья (Магаданская область) минимально необходимый уровень пропуска производителей кеты на нерестилища составляет 1,3—1,4 млн рыб, в Охотском районе — около 2,5 млн рыб. Фактическое изъятие промыслом кеты за последнее 10-летие XX в. составило на северном побережье в среднем 25,5%, в Охотском промысловом районе — 28,7%. Меньшее промысловое изъятие кеты на северо-восточном участке побережья
25
объясняется действием многолетнего запрета на промышленный ловкеты в
реках Тауйской губы в 1970—1980 гг.
Как уже отмечалось, для североохотоморской кеты отмечены два долгопери
одных пика численности на протяжении XX в. синтервалом около 60 лет (рис. 34).")
Рис. 34. Динамика выловакеты на материковом побережье Охотского моря (1920—2009 TT.)
Эти пики численности, проявившиеся и в масштабе всего материкового побережья Охотского моря, в целом совпадают с долгопериодными изменениями численности тихоокеанских лососей, имевшими место в северной Пацифике: первый наблюдался в конце 1930-х гг. второй — в середине 1990-х гг., и оба они согласуются по времени с общим ростом продукции лососей в бассейне Тихого океана (рис. 35).
Максимумы численности лососей в 1936—1938 гг. и в 1994—1996 гг. проявились синхронно как на Азиатском, так и на Североамериканском побережьях Тихого океана.
Суммарные уловы лососей странами Тихоокеанского Кольца в годы пиковых подходов достигли 1 млн т (см. рис. 35), в том числе доля выловакеты составила 47 и 36% соответственно в 1-й и 2-й пики численности. Кроме того, имеются сведения, что подобный пик численности имел место и в 80-х гг. XIX в. (Beamish, Bouillon, 1993; Klyashtorin, Smirnov, 1994). OueBILHo, KeTecBoicTBeHны и более короткие периоды изменения численности, но на наших материалах они явно не просматриваются.
С середины 1990-х гг. и по настоящее время азиатские стада лососей переживают подъем численности. Наиболее четко вспышки численности лососей прослеживаются на Камчатке. Там отмечен значительный рост численностинерки и кеты, их подходы в отдельные годы превышали прогнозные оценки в несколько раз. Так, подходы нерки на Западную Камчатку в 2002 г. превысили прогнозные оценки в 5 раз (Ерохин, 2007б). На североохотоморском побережье косвенным признаком влияния улучшения природного фона в конце XX в. надинамику численности и популяционную структуру кеты может служить фaктроста численности ее ранней формы, которая к 1995 г. достигла 45—48% подходов.
Во втором пятилетии XXI в. возвраты кеты в реки тауйского, ямского и гижигинского стад отмечаются в количествах, близких к 2—2,5 млн рыб. Это
позволяет осуществлять необходимый минимум пропуска производителей на нерести обеспечивать обязательные категории лова (коренных малых народов Севера, научно-исследовательский, спортивно-лицензионный и лов для нужд искусственного воспроизводства). (Klyashtorin, Rukhlov, 1998; NPAFC Statistical Yearbook, 1997-2007)")
Рис. 35. Общий вылов тихоокеанских лососей странами Тихоокеанского Кольца (CIIA, Kahana, AnoHHS, Pocchi) (Klyashtorin, Rukhlov, 1998; NPAFC Statistical Yearbook, 1997-2007)
Теперь попытаемся оценить роль внутривидовых форм кеты в обеспечении промысла. По-видимому, нерестилища LLLLLLLL a LL LLLL LLLL LLLLLLLLS многочисленные нерестовые подходы кеты 1950-х гг., ориентировочная величина которых, вычисленная по уловам, достигала 15—17 млн рыб. Из них 10— 12 млн рыб подходили крекам Охотского района (Клоков, 1970; Костарев, 1970в). Ретроспективный анализ изменений численности и основных показателей биологической структуры охотоморской кеты подтверждает, что до 1960-х гг. основу подходов составляла ранняя кета, воспроизводившаяся на русловых нерестилищах. Лососевый промысел на охотоморском побережье в 1940—1950-е гт. базировался в основном на запасах ранней кеты (Волобуев, Тюрнин, 1995). Об этом косвенно свидетельствует сходный тип динамики численности горбуши и кеты Охотского района в 1945—1955 гг. (Костарев, 1970б). Для кеты в этот периодхарактерно чередование поколений урожайных нечетных и неурожайных четных лет (Костарев, 1964a). В 1940—1950-е гг. в нерестовых подходах преобладала более мелкая кета с биологическими показателями, характерными для ранней формы.
Так, средняя масса охотской кеты в годы массовых подходов 1950—1959 гг. составила 3,25, в последепрессионный период 1971—1984 гг. — 4,09 кг. Нарис. 36 приведено распределение средней массы североохотоморской кеты за последние 45—50 лет для отдельных стад.
Хорошо заметно общее снижение средней массы кеты в 1940—1950-х гг., когда численность ее была наиболее высокой. Наиболее наглядно это прослеживается на кете охотского стада, для остальных стад из-за отсутствия данных имеется четко выраженная тенденция к снижению средней массы в 1950-х гг.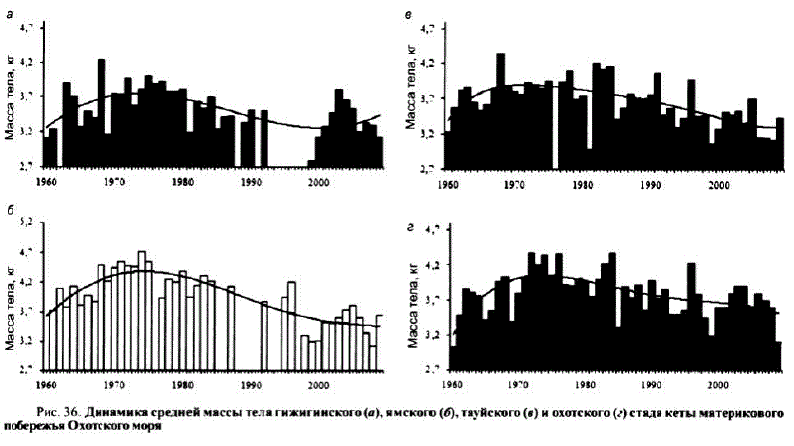
и к середине 1990-х (см. рис. 36). Увеличение средней массы рыб отмечено в период депрессии 1960—1970-х гг., когда численность ранней кеты сократилась почти на два порядка и ее запасы поддерживались в основном благодаря более крупной поздней форме. Очередное снижение средней массы отмечено в 1980— 1990-х гг. в период общего подъема численностилососей в северной Пацифике и возрастания доли ранней формы в подходах североохотоморской кеты к середине 1990-х гг.
Наличие ранней и поздней кеты подтверждается как в реках юго-западного участка побережья (Правдин, 1940; Костарев, 1970в) (охотское и удское локальные стада), так и в его северо-восточной части (Волобуев, 1984, 1986, Волобуев и др., 1990) (тауйское и ямское локальные стада). Ранняя охотоморская кета, как горбуша и летняя амурская кета, размножается в русловой части рек, где икраинкубируется валлювиальном потоке, и в связи с этим она в большей степени, чем поздняя форма, подвержена негативному влиянию климатических условий. Абиотические условия выступают в качестве основного фактора, определяющего ее численность. Резкое сокращение запасов кеты в Охотском рыбопромысловом районе после нескольких неблагоприятных для воспроизводства зим (промерзание и обсыхание русловых нерестилищ) (Костарев, 1964б, 1970б) свидетельствует о том, что численность ранней кеты, как и горбуши, может значительно флуктуировать подвоздействием факторов среды. Поскольку нерестовый фонд ранней кеты гораздо обширнее, чем ее поздней формы, то пристечении благоприятных обстоятельств (оптимальное заполнение нерестилищ, мягкие многоснежные зимы, высокая водность рек, хорошая выживаемость в море) численность ее может нарастать по горбушовому типу, т.е. ранняя экоформакеты является более перспективной для восстановления численности вида в регионе методами искусственного воспроизводства. В связи с этим на материковом побережье Охотского моря необходимо уделять больше внимания искусственному воспроизводству именно ранней кеты.
После 1995 г., когда был достигнут максимум вылова в США (467 тыс. т), североамериканские стада испытали период снижения численности: уловы американских лососей снизились в 1997—1998 гг. и в последующие годы ни разу не превысили максимальный уровень вылова 1995 г. (табл. 41).
В запасах лососей России, напротив, с середины 1990-х гг. отмечается подъем. Наиболее заметный рост наблюдается с 2005 по 2007 г. — с 250 до 350 тыс. т (см. табл. 41). В 2009 г. российский вылов составил 542 тыс. т (с учетом вылова в Исключительной экономической зоне (ИЭЗ) — 551 тыс. т) — это исторически максимальный объем отечественного берегового выловалососей (Гриценко, Кловач, 2010). В целом по Тихоокеанскому бассейну максимальный вылов лососей отмечен в 2007 и 2009 г. — более 1 млн т (см. табл. 41). Такие подъемы и спадычисленности обусловлены долгопериодными циклами изменения глобальных геофизических и климатических характеристик, так называемых режимных сдвигов. Циклические изменения таких факторов, как типы широкомасштабной атмосферной циркуляции, состояние Алеутского минимума и Сибирского максимума атмосферного давления, скорость вращения Земли, температура поверхности океана и др., определяют климатоокеанологическую обстановку в районах основного нагулалососей в северо-западной Пацифике. Конкретные изменения климатоокеанологической обстановки, происходящие с середины 1980-х гг. по настоящее время, заключаются в развитии гидрологических процессов в северо-западной части Тихого океана по холодномутипу. Подвоздействием долгопериодных изменений ветрового режима, температуры и течений происходит изменение общей продуктивности океанических и морских экосистем (Шунтов, 2000). На современном этапе изменение
развития биологических циклов экосистемы северо-западной части Тихого океана, очевидно, привело к сукцессионным процессам в планктонных сообществах, что отразилось и на состоянии кормовой базы лососей. В дальневосточных морях в 1990-е гг. отмечено повышение доли хищного планктона и снижение долимирного планктона, что привело к существенному ухудшению обеспеченности рыб пищей (Шунтов и др., 1998; Шунтов, 2000). В настоящее время биомасса хищного планктона находится на среднем уровне, что свидетельствует об улучшении условий нагулалососей (Шунтов и др., 1998; Старовойтов, 2003).
Таблица 41. Вылов тихоокеанских лососей странами Тихоокеанского бассейна (по данным ежегодных статистических докладов NPAFC), т, т")
Кроме того, под влиянием явления Эль-Ниньо происходят периодические смещения североамериканских лососей на западнагульных акваторий. Значительное потепление за счет Эль-Ниньо северо-восточной части Тихого океана в конце 1990-х гг. оказало негативное влияние на численность массовых видов кормового зоопланктона лососей.
Дефицит кормового зоопланктона усугубляется также появлением на нагульном ареале огромного количества лососей искусственного происхождения - до 5,5 млрд шт. (Кловач, 2003; Heard, 1998). Аналогичных концентраций лососей природного происхождения в местах их традиционных морских пастбиш раньше никогда не было. Избыточность лососей в местах совместного нагула. очевидно, обостряет внутри- и межвидовые конкурентные отношения, включа. ет механизмы плотностно-зависимой регуляции, что выражается в снижении размерно-весовых характеристик, уменьшении продукции отдельных поколений, увеличении возраста созревания, изменении пространственной структуры нагульных скоплений и миграций азиатских и североамериканских стад лососей
(Каев, 1999; Волобуев, Волобуев, 2000; Гриценко и др., 2001; Кловач, 2001, 2003; ATInac., 2002, 3aBapHHa, 2003; Makoenob Hup, 2009; Ishida et al., 1993; Ricker, 1995; Bigler et al. 1996, Helle, Hoffman, 1995, 1998; Kaeriyama, 1996, Volobuev, 2000). Так, за последние 10—12 лет средняя масса тела охотоморской кеты снизилась на 0,3—0,4 кг, длина — на 3—4 см. Увеличение срока созревания рыб в поколениях замедляет скорость смены генераций и приводит к потерям продукции, а кроме того, способствует повышению концентрации рыб в океане (Гриценко и др., 2001).
Результатом последствий массовых выпусков в море заводской кеты и его влияния как фактора плотностной зависимости, по-видимому, может считаться снижение средней массы у анадырской кеты в 1990-е гг. (Макоедов и др., 2009), размерно-весовых характеристик, плодовитости, а также старение популяции кеты о. Итуруп (Каев, 1999, 2003). М.В. Коваль (2009) приводит убедительные данные, свидетельствующие о снижении обеспеченности пищей мощного поколения горбуши 2008 г. северо-востока Камчатки, обусловившем снижение скорости роста и значительное уменьшение ее средних размерновесовых показателей. Есть мнение, что недостаточная кормовая база и переход на питание нетрадиционными объектами питания может приводить к изменениям физиологического состояния и дегенеративным процессам в мышцах кеты (Кловач, 2000, 2001). По мнению автора, увеличение численности одновременно нагуливающихся в море лососей ухудшает условия нагулакеты и в ее рационе возрастает доля низкокалорийных объектов — желетелых организмов (кишечнополостных, оболочников, медуз), что свидетельствует об ограниченности экологической емкости (Саггying capacity) экосистемы северо-западной Пацифики в периоды повышенной численностилососей.
В настоящее время одним из важных факторов, оказывающих влияние на состояние запасов и воспроизводительный потенциал популяций охотоморской кеты, является состояние нерестовых водоемов.
До недавнего времени по долинам ряда основных лососевых рек региона велись интенсивные рубки леса, что вызывало снижение расходов воды в весеннее время — наиболее уязвимый период в раннем онтогенезе лососей (Волобуев, Путивкин, 1986). Кроме того, в бассейнах нерестовых рек проводятся изыскания в целях оценки запасов и передачи в промышленную разработку золото-серебряных месторождений, Планируется добыча бурых углей и разработка месторождений нефти на Примагаданском шельфе. Масштабное развертывание указанных работ неизбежно приведет к значительному сокращению площади и заиливанию нерестилищкеты в рассматриваемом районе. Нефтеразработки планируется проводить вместах весеннего нагула сеголеток лососей, через этот же район пролегают миграционные пути взрослых лососей, идущих на нерест в реки побережья. В связи с этим нельзя исключить негативного воздействия выбросатоксичных буровых растворов и розливов сырой нефти на формирующиеся поколения лососей и реализацию механизмахоминга у рекрутов,
В интересах сохранения естественных мест нереста тихоокеанских лососей считаем недопустимой любую хозяйственную деятельностъ, нарушающую целостность экосистем лососевых водоемов: рубки леса по долинам рек, окультуривание пойменных земель, разработку полезных ископаемых, прокладку коммуникаций и т. д. Практика проведения так называемых компенсационных мероприятий в качестве платы за причиненный ущерблососевому хозяйству малоэффективна и не в состоянии возместить утрату продуктивности природных популяций.