
О НАПРАВЛЕНИЯХ МИГРАЦИЙ ЛОСОСЕЙ РОДА ONCORHYNCHUS В АМУРСКОМ ЛИМАНЕ
Своими знаниями в области пространственного распределения и миграций многих
промысловых гидробионтов мы во многом обязаны многолетней рыбацкой практике.
Однако зачастую мы выстраиваем гипотезы на основе не только рыбацких знаний, но и
рыбацких мифов. Некоторые такие миф-гипотезы могут быть очень живучи. Одной из
них является утверждение о том, что основной путь миграции горбуши в Амур пролега-
ет через пролив Невельского.
В основе этой гипотезы лежит неоспоримый факт, проверенный уже более чем
100-летней историей амурских лососевых промыслов: основная часть горбуши обычно
заходит в устье Амура у правого (южного) берега. В отсутствие других сведений рыба-
кам, а впоследствии и ученым казалось логичным предполагать, что подход к устью
Амура "с юга" связан с миграцией амурской горбуши через пролив Невельского из
Японского моря. Параллельно с этой возникла еще одна миф-гипотеза о "теплолюбиво-
сти" амурской горбуши, предпочитающей идти на нагул прямо на юг (через пролив Не-
вельского), в отличие от амурской кеты, выбирающей для себя путь на север – в Саха-
линский залив и в холодное Охотское море, чтобы затем попасть в океан. Эти гипотезы
еще более окрепли под влиянием данных мечения горбуши (обобщающие публикации:
Takagi et al., 1981; Myers et al., 1990; Атлас распространения… лососей…, 2002; и др.),
убедительно доказывающих, что амурская горбуша действительно нагуливается в Япон-
ском море. Миграция через пролив Невельского в Татарский пролив и Японское море
считается основной для амурской горбуши в работах П.Ю. Шмидта (1905, 1950),
Н.П. Навозова-Лаврова (1927), А.Н. Державина (1933); П.А. Двинина (1959), Р.И. Еню-
тиной (1972) и др.
Вероятно, на основе в том числе и этих миф-гипотез появилась другая, предпола-
гающая, что "теплолюбивая" охотоморская горбуша должна непременно покинуть Охот-
ское море до его охлаждения зимой. Лишь регулярные съемки молоди лососей в Охот-
ском море, проводившиеся ТИНРО (Радченко и др., 1991), развеяли этот миф.
Другая гипотеза, согласно которой горбуша мигрирует в Амур с севера из Охот-
ского моря, была выдвинута В.К. Бражниковым (1900), а впоследствии поддержана
В.К. Солдатовым (1912) и А.Г. Кагановским (1949). Ход горбуши в Амурский лиман и
Амур с севера, пусть и относительно слабый у левого (северного) берега устьевой части
Амура, но зато мощный на северо-западном Сахалине, а также заход кеты (в том числе
осенней) в Амур вдоль правого (южного) берега были сильными доводами в пользу этой
гипотезы. И.Ф. Правдин (1932) также считал эту гипотезу верной, но после получения
данных мечения в Японском море согласился с возможностью прямой миграции части
амурской горбуши через пролив Невельского (Правдин, 1940).
Современные сахалинские исследователи (Иванов и др., 2001) для горбуши северо-
западного Сахалина придают большое значение "южной" миграции, хотя несомненный
приоритет авторы отдают "северной" нерестовой миграции. Вероятно, все же, что авто-
ры имели в виду исключительно горбушу речек Амурского лимана, т.к. сквозная мигра-
ция взрослой горбуши из Японского моря через лиман в Охотское море признавалась
фантастической даже теми исследователями, для которых гидрология лимана Амура
была недостаточно знакома (Бирман, 1985).
И.Б. Бирман (1985) приводит множество косвенных свидетельств возможности
массовой миграции горбуши в Амур через южную часть Охотского моря, в том числе и
из Японского моря. В частности, это данные по миграциям соседних с амурской популя-
ций горбуши (северо-охотского побережья, восточно-сахалинского побережья) в Япон-
ское море. Кроме того, он привел самый убедительный до настоящего времени довод в
пользу гипотезы Бражникова: если бы значительная масса амурской горбуши мигриро-
вала из Японского моря напрямую, она не могла бы остаться незамеченной на промыс-
лах в проливе Невельского. В то же время Бирман не рискнул полностью исключить
возможность миграции какой-то части амурской горбуши этим путем. Немаловажную
роль в этом сыграло, вероятно, отсутствие у него данных о возврате в Амур горбуши, по-
меченной в океане, что оставляло его в плену гипотезы об исключительной "теплолюбиво-
сти" амурской горбуши. И.Б. Бирман акцентирует в своей работе важность получения этой
информации, хотя у К. Такаги с соавторами (Takagi et al., 1981) данные о двух амурских
горбушах, пришедших из океана, уже имелись. Этот бюллетень INPFC (Takagi et al., 1981)
присутствует в списке использованной Бирманом литературы, но, к сожалению, Иосиф
Бенционович, вероятно, имел не полную копию бюллетеня, где эти важные для него
данные отсутствовали. Эти две помеченные в океане амурские горбуши в совокупности
с двенадцатью другими, помеченными в Японском море, уже позволяли утверждать, что
не менее 1/7 части амурской горбуши нагуливается в океане и возвращается в Амур че-
рез Охотское, а не Японское море (как и другие охотоморские и амурские лососи).
В пользу того, что без "южной" миграции амурская горбуша может благополучно
существовать, говорит тот факт, что еще 6 тыс. лет назад пролива Невельского не было
(Гальцев-Безюк, 1964).п
Более полное овладение информацией по мечению, тем не менее, не привело к из-
менению устоявшихся взглядов о морских миграциях амурской горбуши. Так, рис. 1.8 в
издании ВНИРО "Атлас распространения … лососей…" под редакцией О.Ф. Гриценко
(2002. С. 41) за счет формального математического подхода к начертанию областей рас-
пространения лососей вводит в заблуждение читателей о вероятных путях миграции
амурской горбуши. Ежегодно подтверждающийся, известный ученым уже более 100 лет
факт массовых подходов горбуши в Амур со стороны Охотского моря как бы игнориру-
ется. В отличие от этого факта (подчеркиваем – факта, а не гипотезы), миграция амур-
ской горбуши через Татарский пролив и пролив Невельского является лишь гипотезой,
хотя и более часто поддерживаемой в научной литературе. Ни единого факта в пользу
этой гипотезы нет.
Следует отметить, что вопрос о путях миграций амурских и охотоморских лососей
имеет определенное рыбохозяйственное значение в связи с крупномасштабными про-
мыслами в прибрежье практически всего Сахалина, южных Курильских островов и
о-ва Хоккайдо, а также в связи с дрифтерным ловом России и Японии. Например, одни
исследователи утверждают, что ставные невода на Сахалине (длина некоторых из них
достигает 5 км) ловят здесь, за редким исключением, местную горбушу, другие – сомне-
ваются в этом.
Выход «Атласа распространения… лососей…» (2002), реанимирующего устарев-
шую и не подкрепленную фактами гипотезу, явился одной из причин нашего возврата к
результатам своих исследований раннего морского периода жизни тихоокеанских лосо-
сей в Амурском лимане в 1986-1993 гг. Автор настоящей статьи был ответственным ис-
полнителем этих работ. Они проводились под руководством ученика и преемника Вла-
димира Яковлевича Леванидова в Амурском отделении ТИНРО Юрия Сергеевича Рос-
лого. Некоторые полученные нами в Амурском лимане сведения содержатся в его моно-
графии (Рослый, 2002).
Материал и методы
Исследования молоди тихоокеанских лососей в Амурском лимане, Сахалинском
заливе и северной части Татарского пролива проводились на НИС "Волна" (средний
черноморский сейнер, изначально оборудованный как "гидролог"; год постройки –
1951). Орудием лова был 5,4-метровый пелагический трал (Карпенко, Рослый, 1989).
Скорость траления составляла примерно 3,5-4 узла, длина ваеров от 15 до 20 м, горизон-
тальное и вертикальное раскрытие трала примерно 4 м, осадка судна от 2,6 до 2,8 м.
Верхняя подбора и траловые доски во время траления шли у поверхности воды. Про-
должительность тралений варьировала в зависимости от концентрации молоди лососей;
обычно от получаса до часа. Иногда короткие получасовые траления применялись с це-
лью охвата больших акваторий за ограниченное время. По данным сравнения тралового
улова с уловами ловушек на месте проведения учета молоди амурских лососей у
с. Сусанино их коэффициенты уловистости оказались примерно сходными. Коэффици-
ент уловистости ловушек, применяемых Управлением Амуррыброд для учета молоди
лососей в русле Амура, был рассчитан Ю.С. Рослым (1975) по экспериментальным дан-
ным и равен 0,12.
За период с 1986 по 1993 г. было выполнено 405 тралений. Ежегодно исследования
планировались на весь июнь, июль и начало августа, а в 1989 г. Ю.С. Рослым была
предпринята безуспешная попытка обнаружить молодь лососей в сентябре и октябре.
Фактический период и объем работ (табл. 1) определялись в большой степени отрица-
тельными организационными моментами: слабой организованностью финансирования и
снабжения, отсутствием современных навигационных приборов, неприспособленностью
к тралениям при сильном волнении, регулярным выходом из строя механизмов двигате-
ля и пр. К немаловажному положительному моменту организации работ относился ог-
ромный опыт судовождения капитана
Н.И. Кляузова, много лет проработавшего в
Амурском лимане на гидрологических судах.
Существовали и объективные ограничиваю-
щие факторы: так, выставление навигацион-
ных буев на Южном фарватере производи-
лось обычно не раньше 5 июня, на Сахалин-
ском фарватере – не раньше 15 июня, а на
фарватере Невельского – еще позже. Выбор
мест тралений был во многом связан с распо-
ложением судоходных фарватеров (рис. 1).
За весь период исследований нами от-
ловлено более 6 тыс. экз. молоди кеты, более
5 тыс. горбуши и 29 экз. симы. При малых
(<60 экз.) уловах каждого вида измерению и
взвешиванию подвергался весь улов, при
больших уловах – не менее 60 экз.
Использовались также материалы ежегодного учета молоди лососей КНС Амур-
рыбвода в нижнем течении р. Амгунь и в русле Амура у с. Сусанино.
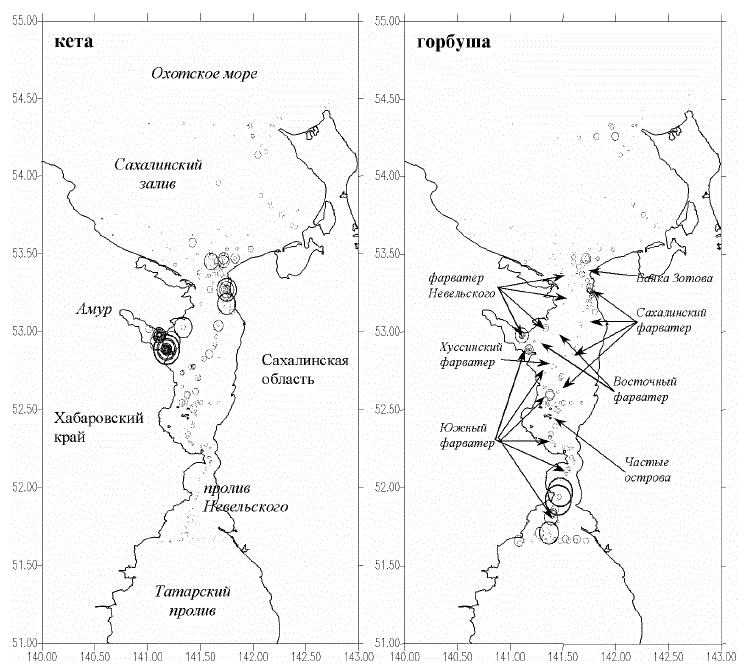
Рис. 1. Район исследований. Судоходные фарватеры. Схема расположения траловых станций и уловы
молоди кеты и горбуши в 1986-1993 гг. (максимальный улов кеты – 1394, горбуши – 1990 экз./ ч траления,
минимальный – 0)
Т а б л и ц а 1
Исследованные периоды и количество
траловых станций
Год Период исследований Количество тралений
1986 25 июля – 16 августа 19
1987 23 июня – 25 июля 56
1988 10 – 22 июня 31
июля – 8 августа 27
1989 30 мая – 20 июля 29
26 сентября – 5 октября 7
1990 30 мая – 23 июля 68
1991 4 июня – 24 июля 59
1992 14 июня – 21 июля 40
1993 2 – 17 июля 32
Результаты и обсуждение
Массовый скат амурской горбуши в Амурский лиман наблюдался нами обычно в
первой декаде июня (рис. 2). К началу третьей декады ее скат прекращался полностью.
Скорость ската лососей по Амуру примерно равна скорости течения (Рослый, 1975), по-
этому мальки от мест учета в Амгуни и у с. Сусанино доходят до устья Амура примерно
за 2 сут. Исходя из этого, по данным Амуррыбвода (рис. 3) видно, что массовый выход
горбуши в Амурский лиман может начинаться во второй половине третьей декады мая, а
всплески массовой миграции могут быть и на 10 сут раньше. Сопоставление данных
(рис. 2 и 3) указывает на то, что большую часть амурской горбуши, а в 1989 г. – всю мы
своими исследованиями в лимане Амура охватывали, и появление в южной части Амур-
ского лимана относительно большого количества амурской горбуши было бы нами за-
мечено. Однако июньские концентрации горбуши к югу от Частых островов и в проливе
Невельского были всегда меньше, чем в устье Амура и в северной части Амурского ли-
мана (у Рыбновска и банки Зотова), а также в южной части Сахалинского залива (рис. 1).
В среднем июньские уловы горбуши там были ниже, чем в устье Амура, примерно в 5
раз, и ниже, чем в районе банки Зотова, примерно в 2 раза (табл. 2).
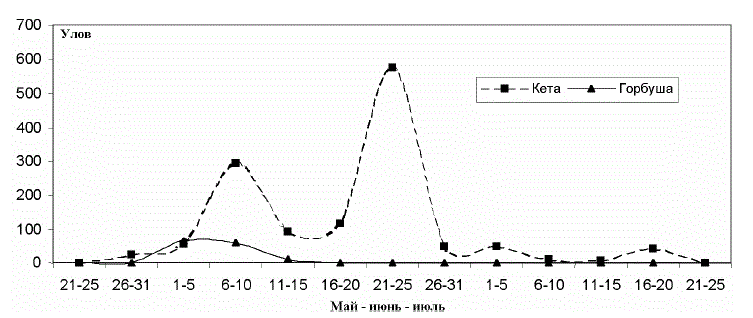
Рис. 2. Усредненные уловы в устьевой части Амура в 1998-1993 гг., экз./ ч траления Т а б л и ц а 2
Т а б л и ц а 2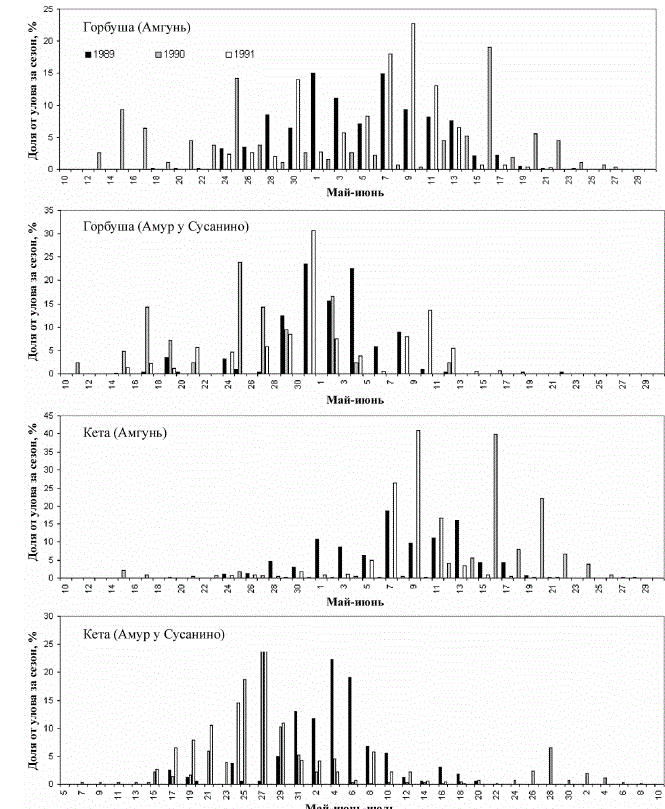
Рис. 3. Динамика ската молоди лососей в контрольных точках КНС Амуррыбвода в 1989-1991 гг.
В северной же части Татарского пролива и на входе в пролив Невельского как в
июне, так и июле отмечались огромные (по сравнению с уловами в устье Амура и в ли-
мане) уловы горбуши (от 1000 до 2000 экз. на час траления) при практически полном
отсутствии кеты (рис. 1, табл. 2). Стаи мальков с примерно таким же соотношением гор-
буши и кеты мы визуально наблюдали в это время в порту поселка Де-Кастри недалеко
от устья р. Сомон. В стае, состоящей из нескольких сотен или даже тысяч мальков гор-
буши, присутствовали от 1 до 3 мальков кеты. Горбуша на входе в пролив Невельского
в июле была в среднем мельче, чем в южной части лимана (рис. 4, з, и), что не укладыва-
лось в логику "роста ло-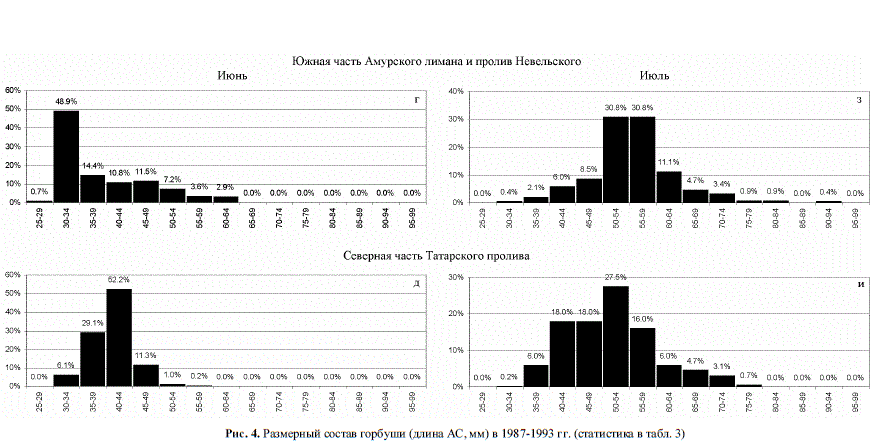
сосей по мере удаления от
реки", в отличие от Саха-
линского залива, где такая
закономерность обычно
проявлялась (рис. 4,в→
4,б, 4,ж → 4,е, 6,в→ 6,б→
6,а, 6,ж→ 6,е→ 6,д). Эти
данные указывали, что
облавливаемая в Татар-
ском проливе горбуша не
из Амура и даже не из
южной части Амурского
лимана.
На то, что более вы-
сокие уловы в северной
части Татарского пролива
связаны с япономорским
происхождением этой
горбуши, косвенно ука-
зывает также динамика
вылова северо-примор-
ской и амурской горбуши
(рис. 5). В период наших
исследований северо-
приморская горбуша обо-
их поколений была на
подъеме численности, а в
нечетные годы проходила
период максимума. Амур-
ская же горбуша нечет-
ных лет в эти годы была в
депрессии, а в четные
годы ее численность была
не самой высокой для
этих поколений.
Сопоставляя выше-
перечисленные сведения
по горбуше со сведения-
ми по кете (рис. 1, 2, 3, 6,
табл. 2), мы видим, что
молодь этих видов в
Амурском лимане мигри-
рует совершенно одина-
ково – на север – в Саха-
линский залив и далее.
Кета, так же как и горбу-
ша (до половины своей
численности), может ска-
титься в лиман уже к кон-
цу мая (рис. 2 и 3), однако
мы не находили ее в
большом количестве ни в
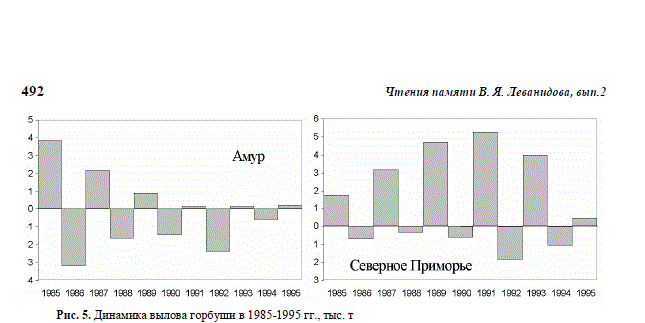
Рис. 4. Размерный состав горбуши (длина АС, мм) в 1987-1993 гг. (статистика в табл. 3)
южной части Амурского лимана, ни, тем более, в северной части Татарского пролива
(рис. 1).
А.Г. Поддубный и Л.К. Малинин (1988) считают, что крейсерская скорость рыб (в
том числе лососей) редко превышает 1 длину рыбы в секунду. Д.С. Павлов (1979) по
данным Блэкстера (Blaxter, 1967) полагает, что у лососей она примерно 3-4 длины рыбы
в секунду. Минимальные скорости непериодических течений на фарватерах Амурского
лимана в зоне влияния Амура редко бывают менее 10 см/с, а максимальные достигают
50 см/с и выше (Якунин и др., 1977; Соловьев, 1995). Получается, что скорости течений
в исследуемом районе обычно превышают крейсерскую скорость мальков не только
горбуши, но и кеты (рис. 4, 6). Представляется весьма сомнительным, чтобы мальки гор-
буши, мигрирующие по фарватеру, могли каким-то образом сильнее влиять на свое
дальнейшее направление миграции, нежели мальки кеты. Последних течение выносит в
район банки Зотова (рис. 1), причем независимо, каким фарватером мальки вышли в ли-
ман – северным или южным. Попадание молоди лососей из Южного фарватера в район
банки Зотова вполне объяснимо, т.к. именно так ведут себя воды этого фарватера (Яку-
нин и др., 1977). После выхода из Амура течение Южного фарватера поворачивает на
юг – юго-восток, постепенно веерообразно отклоняется на восток и сливается с течени-
ем Сахалинского фарватера, идущего на север. Течение Южного фарватера сохраняет
свою мощь до Хуссинского фарватера, но в районе Частых островов влияния Амурского
течения уже не ощущается. Течение Южного фарватера на участке от м. Пронге до
м. Джаорэ является единственным непериодическим течением Амурского лимана, на-
правленным с севера на юг. Все остальные непериодические течения, в том числе в про-
ливе Невельского, на протяжении всего года направлены на север (Якунин и др., 1977).
Предпочтение молодью горбуши Южного фарватера (при выходе из Амура) с це-
лью дальнейшей миграции "на юг" вытекает из логики исследователей, проповедующих
гипотезу миграций амурской горбуши через пролив Невельского. Однако в период на-
ших исследований это не подтвердилось. Так, средние уловы молоди горбуши в период
с 1 по 20 июня (1989-1992 гг.) в устье Амура на фарватере Невельского составляли
38,3 экз./ч траления (mах=454, n=20), а на Южном фарватере – 31,0 экз./ч (mах=164,
n=14). Северный путь миграции у горбуши даже преобладал: как в среднем, так и по
максимальным уловам. Уловы кеты за аналогичный период в северном и южном рукаве
оказались практически равными: 110,1 экз./ч траления (mах=632) и 102,0 (mах=531).
Кстати, большое количество молоди лососей выходит в Амурский лиман посредине
устья – через Восточный фарватер. 24 июня 1990 г. мы провели траления на всех трех
рукавах устьевой части Амура. Улов на фарватере Невельского составил 96 мальков ке-
ты за час траления, на Южном фарватере – 711, а на Восточном фарватере – 546 кеты и
две горбуши. Мальки горбуши вероятно были из р. Амгунь, т.к. у Сусанино мальки гор-
буши в это время уже не скатывались (рис. 3).
Таким образом, механизм выбора амурскими лососями направлений и путей ми-
граций в ранний морской период пассивен, случаен, связан с направлениями течений и
Рис. 5. Динамика вылова горбуши в 1985-1995 гг., тыс. т
Г.В. Новомодный 493
одинаков у кеты и горбуши. Такой же механизм, несомненно, присущ и лососям из дру-
гих рек Амурского лимана. Логично предполагать, что нерестовая миграция лососей
зеркальна относительно миграции в ранний период жизни. Поэтому представляется со-
мнительной возможность миграции горбуши в реки северо-западного Сахалина через
пролив Невельского (Иванов и др., 2002). В этом случае ей пришлось бы двигаться по
течению, и реализация реотактильно-обонятельного механизма ориентации (Dodson,
Leggett, 1974), присущего лососям во время нерестовой миграции (Поддубный, Мали-
нин, 1988), была бы не возможна.
Напротив, мы полагаем, и у нас есть косвенные подтверждения того, что из Охот-
ского моря через Амурский лиман и пролив Невельского вдоль сахалинского побережья
в Татарский пролив происходит нерестовая миграция горбуши. Косвенными подтвер-
ждениями этому являются данные о направлениях миграций молоди лососей, а также
данные гидрологии в Амурском лимане:
• непериодическое течение, постоянно направленное из Татарского пролива в Амур-
ский лиман (Юрасов, Яричин, 1991), и размерный состав молоди горбуши в север-
ной части Татарского пролива и в южной части лимана (рис.4,з,и), указывающий на
миграцию "в лиман", а не из него;
• мощные периодические сквозные течения через Амурский лиман из Татарского
пролива в Сахалинский залив под действием штормовых ветров в совокупности со
снижением стока р. Амур, ежегодно наблюдаемые нами в конце июня и июле;
А.К. Леонов (1960) считал причиной этого также усиление Куросио и его ветви –
Цусимского течения, создающего подпор вод в Татарском проливе;
• эти июльские сквозные течения хорошо просматриваются по схемам распределе-
ния солености в Амурском лимане (Юрасов, Яричин, 1991); по нашим данным, в
июле соленость около Частых островов может достигать 30 ‰;
• факты переноса япономорских вод в Сахалинский залив фиксировались нами также
и по биологическим маркерам; в частности, в Сахалинском заливе в июле появля-
лась молодь южного одноперого терпуга, отсутствующая здесь в июне; в июне мо-
лодь этого вида в массе облавливалась вместе с горбушей в северной части Татар-
ского пролива;
• обнаружение в июле в районе Частых островов молоди горбуши с присущим Та-
тарскому проливу размерным составом и без характерного для южной части лима-
на относительно большого прилова кеты.
Генерализованная схема миграций молоди лососей в Амурском лимане и приле-
гающих к нему акваториях (рис. 7) составлена нами исходя из представлений о не-
периодических течениях в данном районе (Юрасов, Яричин, 1991). Эта схема уже
ранее представлена нами в NPАFC как схема миграций молоди лососей и их основ-
ного хищника в Амурском лимане и Сахалинском заливе – молоди миноги (Novomodnyy,
Belyaev, 2002).
Несмотря на сходство путей и направлений миграций, у мальков горбуши и кеты в
Амурском лимане наблюдается несколько различная стратегия образования "ранних" и
"поздних" мигрантов. "Поздние мигранты" – это дольше задерживающиеся для нагула в
реке и морском прибрежье мальки; "ранние" – быстро скатывающиеся из реки и рано
переходящие к пелагическому образу жизни в море (Рослый, Новомодный, 1991; Рос-
лый, 2002; и др.). Различная миграционная стратегия видов связана с определенным,
отличающимся у них набором врожденных поведенческих реакций: предпочтение того
или иного корма, та или иная реакция на течение, на свет, на температуру и соленость,
на глубину, на типы грунтов, на хищников, на собратьев и пр. (Павлов, 1979; и др.).
Исходя из полученных нами данных, видно, что стратегия горбуши – раньше (не
задерживаясь, как кета) скатиться из рек (рис. 2, 3), при этом сильно уступая кете в раз-
мерах (рис. 4,в, г, 6,в, г), занять мелководное, слабо осолоненное прибрежье, в частно-
сти, южную часть Амурского лимана, где условия для ее быстрого роста очень благо-
приятны (рис. 4,г). Амурской горбуши здесь не много или вообще нет, т.к. амурское те-
чение в этот район уже практиче-
ски не доходит. Большая часть
горбуши здесь из речек южной
части лимана (Мы, Уаркэ, Чомэ,
Тыми), пролива Невельского
(Псю и Нигирь) и из Татарского
пролива.
Уже к концу первого месяца
жизни в лимане размеры горбу-
ши существенно превышают
размеры кеты, проведшей такое
же время в пресных водах
(рис. 4,е, ж, з, 6,ж). Приустьевое
мелководье является первым ру-
бежом, где формируется сущест-
венная доля "поздних мигрантов"
горбуши.
Нельзя полностью отрицать
возможность образования "позд-
них мигрантов" у горбуши в реке,
так как мы обнаружили 3 малька
горбуши с размерами более 50 мм
в сборах Амуррыбвода из русла
Амура у Сусанино за начало июля
1983 г. В принципе, это лишь
подтверждает данные Енютиной
(1972) о сроках ската из нересто-
вых рек Им и Сомня (бассейн
р. Амгунь; окончание массового
ската – 16 июня, полное оконча-
ние – 20 июля–2 августа). Однако
таких крупных мальков горбуши
в ее распоряжении не было. По
данным Р.И. Енютиной (1972),
размер июльских покатников в
реках Им и Сомня практически не
отличается от майских и июнь-
ских. "Поздние речные мигранты"
горбуши в Амуре обычно поги-
бают из-за высокой температуры
в июле, т.к. по нашим данным их
нет в устье Амура (рис. 2 и пустое
место между рис. 4,ж и 4,з). По
данным Р.И. Енютиной (1972;
р. Мы) "поздние речные мигран-
ты" в южно-лиманных речках так-
же не крупные – менее 43 мм.
Условия для их выживания, в от-
личие от Амура, имеются, но их
доля в общей массе мальков гор-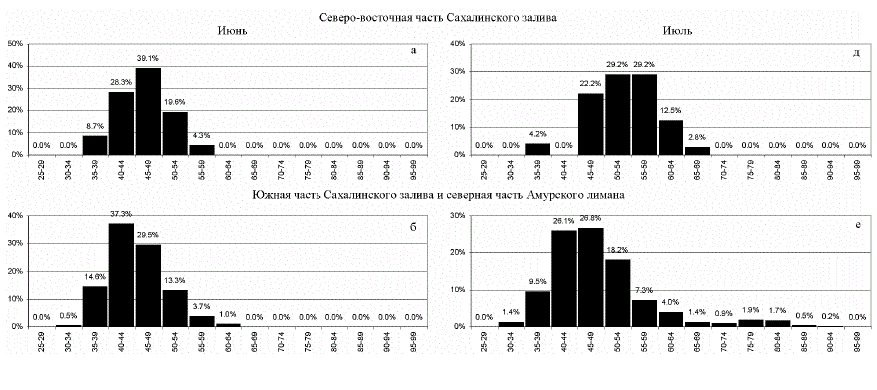
буши, тем не менее, чрезвычайно мала (рис. 4,з).
И все же большая часть горбуши, как и большая часть кеты, при выходе из Амура
не попадает на мелководье и по фарватерам сразу же уносится в море (рис. 4,а, б, 6,а, б),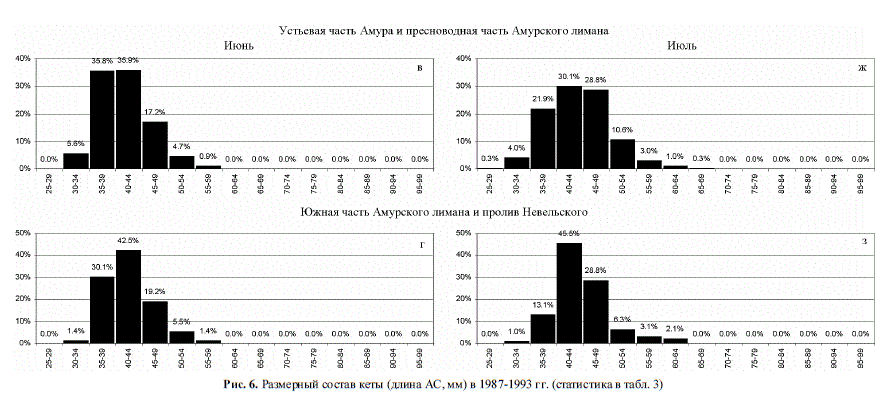
превращаясь в так называе-
мых ранних мигрантов. Ско-
рость миграции молоди лосо-
сей в пелагиали Сахалинского
залива практически равна
скорости течения. Это видно
по "приросту" молоди между
южной и северной оконечно-
стью Сахалинского залива
(рис. 4,б и 4,а, 4,ж и 4,е, 6,б и
6,а, 6,е и 6,д). Прирост соот-
ветствует 5-10-суточному из
расчета 0,5-1 мм/сут. 5-10 сут
вполне достаточно, чтобы
преодолеть 100 км со скоро-
стью 10-20 см/с. Молодь раз-
ная по размеру, при этом ско-
рость миграции у них одина-
ковая. В период проведения
исследований у нас появилась
гипотеза о механизме нагуль-
ной миграции лососей (Ново-
модный, 1992). При относи-
тельно равномерном распре-
делении корма в пределах
досягаемости органов чувств
лосося, вследствие чего нача-
ло движения к корму тем или
иным направлением равнове-
роятно, значение суммарного
вектора активного перемеще-
ния стремится к нулевому, а
значит вектор перемещения
лосося в пространстве стано-
вится равен вектору неперио-
дического течения.
Следующим рубежом,
где происходят задержка мо-
лоди и образование "поздних
мигрантов" в Амурском ли-
мане, является мелководный
участок в районе Рыбновска,
банки Зотова и банки Север-
ная. Это мелководье уже бо-
лее осолоненное, чем район
Частых островов за счет регу-
лярных приливов в этот район
с северо-запада. В отличие от
мелководий южной части ли-
мана, где кета практически не
задерживается, что видно из данных по ее размерному составу в июне и июле (рис. 6,г,
з), в районе банки Зотова условия для формирования морских "поздних мигрантов" кеты
имеются (рис. 6,е). Здесь же формируется и большая доля "поздних мигрантов" горбуши
Рис. 6. Размерный состав кеты (длина АС, мм) в 1987-1993 гг. (статистика в табл. 3)
(рис. 4,б, ж). В этом районе облавлива-
ется смесь молоди лососей различного
происхождения (амурского, южно-ли-
манного сахалинского и южно-лиман-
ного материкового, япономорского). За
счет приливов с северо-запада сюда,
вероятно, попадает молодь из залива
Счастья (р. Иска).
В северной части Сахалинского за-
лива "поздние мигранты" кеты не об-
лавливаются (рис. 6,д). В уловах горбу-
ши здесь "поздних мигрантов" также
существенно меньше, чем в южной час-
ти Сахалинского залива (рис. 6,е, ж).
Вероятно, это может быть следствием
контакта с молодью миноги, выедающей
и смертельно травмирующей мальков
лососей. Известно, что в пелагиали се-
верной части Сахалинского залива
мальки лососей и минога остаются
практически один на один друг с другом
(Рослый, Новомодный, 1986). При раз-
боре улова складывается впечатление,
что имеет место селективность выеда-
ния миногой более крупных мальков
лососей в северной части Сахалинского
залива. Статистического подтверждения
этому пока нет (Рослый, Новомодный,
1996): травмированная и нетронутая
миногой молодь из наших уловов досто-
верно не различалась.
Мелководье в южной части Саха-
линского залива и сахалинского при-
брежья Амурского лимана является ме-
стом длительной задержки не только
молоди горбуши и кеты, но и симы. В
этом районе поймано 23 экз. из 29 экз.
общего улова симы тралом за период с 1986 по 1993 г. Кроме того, сима нередко облав-
ливалась нами мальковым неводом в заливе Счастья и на банке Зотова в июле. Встре-
чающаяся в Амурском лимане и соседних участках морей в июле молодь симы имеет
длину от 98 до 160 мм (табл. 3, рис. 8). Вероятно, сима держится здесь до сентября, т.к. у
нас имеются сведения о массовом попадании в заездок (разновидность ставного невода)
у поселка Рыбновск каких-то молодых лососей, называемых рыбаками "горбуша".
Имеющиеся в нашем распоряжении фотографии не позволяют точно определить их вид.
Это может быть также и молодь кижуча. В нашей коллекции имеются 2 кижуча, пой-
манных сотрудником ХоТИНРО С.С. Юхименко неводом в южной части Сахалинского
залива (Петровская коса) в начале сентября 1990 г. Их длина аналогична длине молоди
из Рыбновска – около 25 см. Обнаружение молоди кижуча в районе Петровской косы не
случайно. В последнее время нами выяснено, что кижуч является относительно мало-
численным, но обычным приловом к кете в материковых реках Сахалинского залива
(Иска, Коль, Тывлина).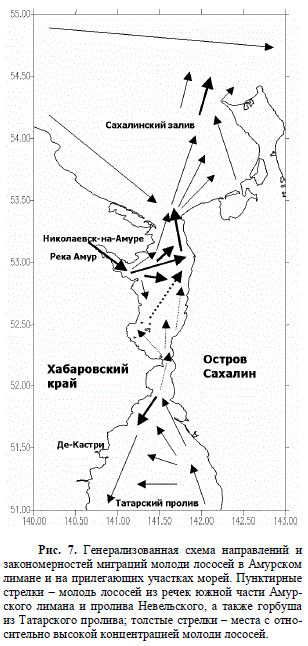
Рис. 7. Генерализованная схема направлений и
закономерностей миграций молоди лососей в Амурском
лимане и на прилегающих участках морей. Пунктирные
стрелки – молодь лососей из речек южной части Амур-
ского лимана и пролива Невельского, а также горбуша
из Татарского пролива; толстые стрелки – места с отно-
сительно высокой концентрацией молоди лососей.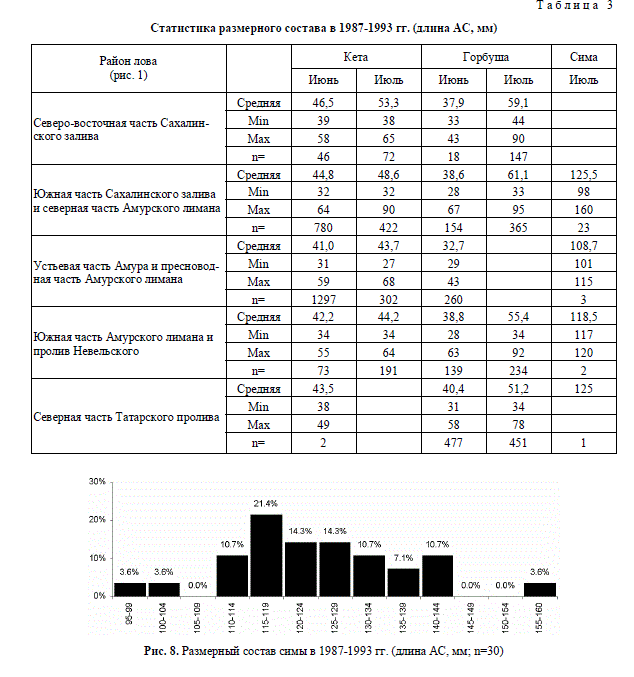
Рис. 8. Размерный состав симы в 1987-1993 гг. (длина АС, мм; n=30)
Выводы
Миграции молоди амурских лососей через пролив Невельского нет, соответственно
нет и обратной нерестовой миграции. Амурская горбуша попадает в Японское море так
же, как и соседние популяции (северо-охотоморские и восточно-сахалинские), – через
пролив Лаперуза, а в океан так же, как амурская кета, – через южные Курильские про-
ливы.
Вероятность миграции через пролив Невельского для молоди из речек южной час-
ти лимана и пролива Невельского низка. Соответственно и нерестовая миграция в об-
ратном направлении маловероятна.
Через Амурский лиман из Татарского пролива в Сахалинский залив происходит
транзит молоди горбуши. Должна так же существовать и обратная нерестовая миграция
горбуши с севера вдоль сахалинского берега Амурского лимана в Татарский пролив.
Направления миграций молоди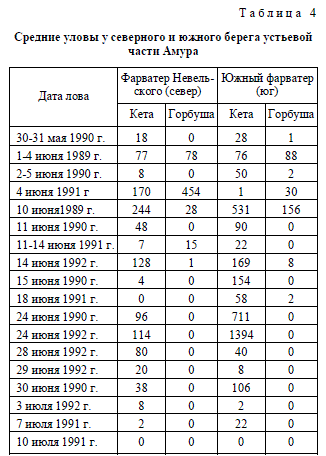
лососей в Амурском лимане и на при-
легающих к нему участках морей аб-
солютно соответствуют направлениям
течений. Схему течений и миграций
молоди лососей в Амурском лимане
можно описать в общих чертах, как
"из пролива Невельского и Амура в
Сахалинский залив".
Немалая часть амурских и дру-
гих лососей образует перед уходом в
море скопления так называемых
поздних мигрантов в северной части
Амурского лимана и южной части
Сахалинского залива (район Рыбнов-
ска и банок Зотова и Северная). Здесь
же длительное время (июль–август;
вероятно до сентября) нагуливается
сима. В сентябре у берега Сахалин-
ского залива облавливается подрос-
шая до 25 см молодь кижуча.
У кеты большая часть "поздних
мигрантов" формируется в пресных
водах, а также на достаточно сильно
осолоненных мелководьях прибрежья.
У горбуши "поздних речных мигран-
тов" мало или, как в Амуре, практиче-
ски нет. "Поздние речные мигранты" горбуши обычно не на много крупнее "ранних ми-
грантов". Горбуша в большом количестве задерживается на 1-1,5 мес как в слабо осоло-
ненном прибрежье, которое кета избегает, так и в сильно осолоненном прибрежье. Раз-
меры "поздних мигрантов" горбуши перед уходом в море достигают 95 мм, кеты – 90 мм.
Г.В. Новомодный
Хабаровское отделение ТИНРО-Центра, ул.Шевченко, 9,
Хабаровск, 680000, Россия. E-mail: Этот адрес электронной почты защищён от спам-ботов. У вас должен быть включен JavaScript для просмотра.
Смотрите также