
Эмбриональный, или зародышевый, период развития чавычи
1. Обводнение икры, образование зародышевого диска. Сбор и искусственное оплодотворение икры проведены в районе нерестилища. Перевозку икры осуществляли в изотермическом ящике. Икру доставляли в лабораторию и помещали в инкубатор через 31 и 52 часа после осеменения. При более быстрой перевозке икра до закладки в инкубатор достигала стадии 32 бластомеров.
По данным Риддла (Riddle, 1917), развивающаяся икра чавычи имеет диаметр 7—9 мм (среднее 7,5 мм) и вес 260—330 мг. Прескотт (Prescott, 1955) сообщает, что вес икринок в результате обводнения возрастает на 15,5%, а объем — на 16,5%. В опытах с тяжелой водой (D2O) он убедительно доказал образование перивителлиновой жидкости за счет поступления в яйцо воды из внешней среды. Вес икринок разных самок из р. Паратунки после набухания увеличивался от 4,7 до 13,6%, их диаметр равнялся 7,5— 7,7 мм, средний вес 227—258 мг (вес отдельных икринок достигал
267 мг). Было замечено, что меньше воды воспринималата нкра, которая дольше задерживалась в теле самки.
Во время набухания оболочка икры становится полупрозрачной, плотной, эластичной. Ее толщина, по данным Риддла, 64,7 мк. У набухших икринок камчатской чавычи толщина оболочек колебалась от 380 до 57,0 мк (53,75+0,78). Набухшие икринки покрывались белесым глянцеватым налетом, дающим в отраженном свете характерныи перламутровый блеск. Микропилярное поле, имеющее диаметр немного более 1 мм, лишено такого HarleTa.
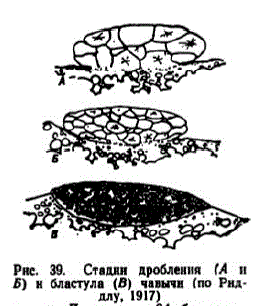
2 этап. Дробление заоодышевого диска. Первая бороздка дробления формировалась примерно через 4,5 граIW'C0-III H I10CIe 0ceMeHeHHRH. Последующие дробления наступали через 5,2-55, 6,3- 6,6, 7,4-77, 8,5-8.8, 9.7-10.0 и 10,8—11,2 градусо-дней. Борозды дробления располагаKAK HI ???? лососей. о Риддлу (1917), при пятом дроблении борозды проходят ?°, ac? '' в экваториальной плоскостин длу, 1917) образуют зачаток из двух слоев клеток. До стадии 64 бластомеров зародышевый диск сохраняет немного вытянутую форму, а со стадии 128 бластомеров становится округлым.
На этой стадии между нижележащими бластомерами образуются межклеточные промежутки, которые в последующем, по мнению Риддла, дают полость между бластодермой и перибластом (рис. 39).
На нашем материале в экваториальной плоскости проходили борозды 4-го дробления, образуя 2 слоя клеток (рис. 40). Первая борозда проходила глубже других и ее расположение можно было выявить до образования 64 бластомеров. После стадии 128 клеток (в инкубаторе в возрасте 1 сут. 22 час.) бластодиск приобретал округлую форму с днаметром около 1 мм. До стадии 128 бластомеров чавыча развивается при температуре не ниже 4,4—5,8°. Верхний температурный порог лежит в пределах 14,2—15,5° (Combs, Burrows, 1957; Combs, 1965).
В возрасте 3 сут. (21 градусо-день) бластодиск имел диаметр 1,0—1,2 мм и высоту 0,45—0,49 мм. Диаметр клеток поверхностного слоя был равен 75—80 мк. Позже более мелкие и плотно прилегающие друг к другу клетки покрывали массу свободно расположенных клеток. Снизу бластодиск подстилал перибласт; при температуре 7,5-12 начало его формирования отмечено через 44 час. после осеменения.
При 4°тысяча икринок в возрасте 4 сут. потребляла за 1 час 1,41 мг кислорода (среднее по двум определениям). Эта величина меньше по сравнению с потреблением кислорода во время набухания икры. Позже интенсивность обмена постепенно возрастает (табл. 13).
Таблнца 13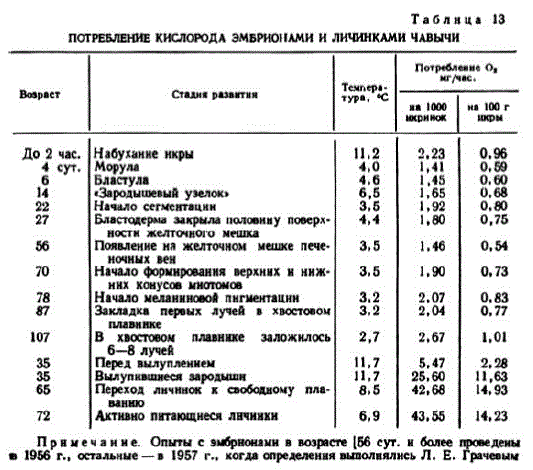
Прн меч анне. Опыты с эмбрнонамн ввозрасте (56 сут. н более JeHIb
в 1956 г., остальные — в 1957 г., когда определения выполнялись Л. Е. Грачевым
3 этап. Бластула. В возрасте 5 сут. 9 час. зародышевый диск имел вид, сходный с описанным Риддлом через 101 час после оплодотворения (рис. 39, В). Его диаметр увеличился до 1,35—1,5 мм. Диаметр клеток покровного слоя бластулы уменьшился до 40 мк (рис. 40, E; 26—28 градусо-дней). Со временем бластодиск расширялся и уплотнялся. В первой стадии этап длился около 5,5 сут., во второй — немного более суток.
4 этап. Гаструляция. При температуре 3,7—4,1° признаки гаструляции отмечены в начале 11 сут., при 15,5—16,5° — в середине 4 сут. Зародышевый диск имел диаметр 1,7—2,1 мм. Диаметр клеток поверхностного слоя уменьшился до 20—25 мк. Через утоньшенную центральную часть диска просматривались лежащие снизу мелкие жировые капли. С каудальной стороны закладываю
щегося зародыша краевой участок диска начал утолщаться (рис. 40, Ж).
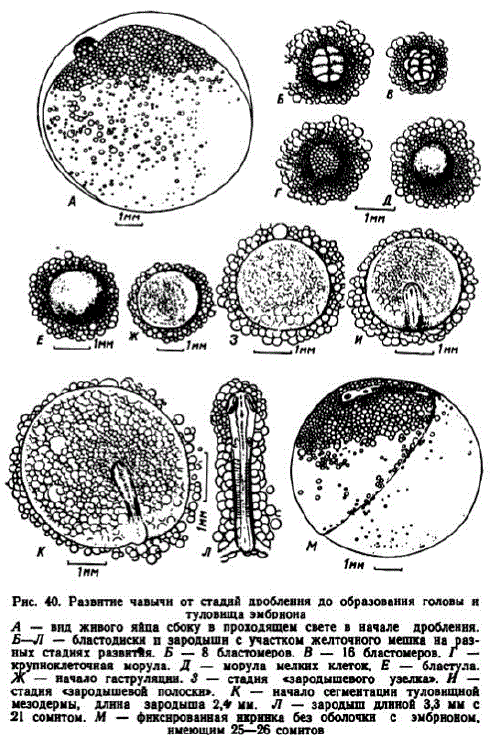
С увеличением диаметра бластодиска до 2,5-3,2 мм (68 градусо-дней) образовался «краевой узелок» (рис. 40, 3). Границы эмбрионального щита вышли за зону скопления мелких жировых капель. В речных аппаратах в возрасте 5 сут. 5 час. (около 75 градусо-дней ????????????- зародышевая полоска длиной 1,3—1,4 мм (рис. 40, И) началом гаструляции икринки стали очень чувствителih himih k toj likem.
5 этап. Формирование головы и туловища зародыша. В икринках первой серии сегменты появились в возрасте 19—20 сут. (80—84 градусо-дня), во второй — в начале 6-х сут. К этому времени диаметр бластодиска достигал 3,5—4 мм, а длина зародышей — 2—2,2 мм. Некоторые эмбрионы в речных аппаратах в возрасте 5 сут. 17 час. имели по 3—4 сегмента; у них наметилась дифференцировка отделов головного мозга (рис. 40, K).
В инкубаторе через 2 сут. после начала сегментации эмбрионы достигали длины 3,2 мм. У них сформировалось по 7—8 сомитов, обозначились зачатки глаз. Через сутки, на стадии 12—13 сомитов, появились зачатки хрусталиков и слуховые пузырьки. Когда у эмбрионов насчитывалось 20—22 сегмента (около 100 градусодней), головной отдел составлял 34,5% их длины (рис. 40, Л).
В возрасте 25 сут. (104 градусо-дня) бластодерма охватывала до половины сферы желточного мешка. Зародыши достигали длины 3,3—3,6 мм, у них насчитывалось 25—26 сегментов. Образовалась перикардиальная полость, в которой размещался зачаток сердца. Основная массажировых капель сосредоточивалась вблизи тела эмбриона и лишь немногие более мелкие находились за пределами зоны обрастания (рис. 40, M).
К 34 сут. (138 градусо-дней) бластодерма покрывала */s поверхности желточного мешка или немного большую часть. В икре из речных аппаратов сходная стадия наблюдалась в возрасте 8 сут. (115 градусо-дней). Эмбрионы достигли длины 4,1—4,4 мм и у них насчитывалось 35—37 сегментов (рис. 41, A). Образовалась небольшая хвостовая почка. Размер головного отдела сократился до 28% длины зародыша. Слуховые капсулы от глаз были удалены на расстоянне, почти равное ширине 10 сегментов. Относительно небольшие глаза имели диаметр около 0,4 мм. Обозначились обоиятельные плакоды. Четко выявились отделы головного мозга и энцефаломеры. Когда бластодерма покрыла большую половину желточного мешка, чувствительность икринок к толчкам на некоторое время снизилась.
6 этап. Обособление задней части тела зародыша от поверхности желточного мешка. В конце 36 — начале 37 сут. (145— 150 градусо-дней) зародыши достигли длины 4,9—5,2 мм и у них сформировалось 45—48 сегментов. Сохранялась лишь небольшая желточная пробка. Продолжала дифференцироваться хвостовая
почка. Появились зачатки жаберных крышек. В этом состоянии наблюдались слабые сокращения миотомов. Сердечная трубка приняла изогнутую форму и у эмбрионов сббльшим числом миотомов волнообразно сокращалась. При развитии в речных аппаратах пульсация сердца отмечена в возрасте 9 сут., по прошествии всего лишь 129 градусо-дней. В этом возрасте чувствительность икры к толчкам резко возрастала.
В мелких икринках первой серии процесс эпиболии завершался у эмбрионов длиной 5,5—6,0 мм на стадии 48—49 сегментов (развитие с 5/VIII по 14/IX 1956 г., около 160 градусо-дней). В это время длина головы составляла 23% длины эмбрионов. От желточного мешочка отделилась хвостовая почка, равная ширине семи туловищных миотомов. Ниже 43—44 сомитов сформировалось анальное отверстие. Появился зачаток непарной плавниковой складки. У основной массы зародышей эпиболия завершалась при 53—56 сегментах, а в крупных икринках — после образования 58 и 59 сегментов. При более высокой температуре у отдельных икринок обрастание желточного мешка бластодермой продолжалось достадии 63—64 сегментов. Таким образом, в зависимости от качества икры (ее размеров) и условий инкубации процесс эпиболии заканчивается на разных стадиях морфологической дифференцировки. Имеется указание на завершение этого процесса у эмбрионов североамериканской чавычи длиной 4 мм, имевших 38 сегментов (Riddle, 1917). Такое ускорение эпиболии, возможно, свидетельствует о генетических особенностях материала, который был в распоряжении Риддла. Во время завершения эпиболии икра очень чувствительна к толчкам.
7 этап. Развитие подкишечно-желточной системы кровообращения. В условиях низкой температуры циркуляция крови у разных эмбрионов обнаруживалась через 43—47 сут. после оплодотворения (172—187 градусо-дней), когда они достигали длины 7—7,5 мм и имели по 64—66 сегментов. К этому времени хвост, в котором насчитывалось 21—22 сегмента, и задний отдел туловища обособились от желточного мешка. Голова по-прежнему плотно прилегала к желточному мешку, а ее длина продолжала уменьшаться. Появились зачатки жаберных крышек. Начали формироваться обонятельные ямки. Слуховые пузырьки заметно увеличились; в них дифференцировались полукружные каналы и были видны многочисленные мелкие отолитики. От 15 или 17-го миотома вдоль спины тянулась узкая плавниковая складка. Значительной длины достигла преанальная складка. Заложились грудные плавнички. Позади них просматривались клубочки пронефроса. Наметилось обособление желудка 1.
1. Автор не нмел возможности поместить подготовленные им рисунки по развитню желудочно-кншечного тракта, плавательного пузырян жабер чавычн н кнжуча.
В начале этапа форменных элементов в сосудах мало, эритроциты окрашены слабо, кровь течет медленно и омывает лишь два сегмента хвостового отдела. Схема кровообращения сходна с описанной у кеты (рис. 41, Б).
У эмбрионов, развивающихся в речных аппаратах, кровообращение начиналось при 63—64 сегментах (101/4 сут., 147 градусодней). У некоторых из них оставалась небольшая желточная пробка. Эмбрионы периодически наклонялись то на правый, то на левый бок и медленно поводили хвостами. При температуре около 14,5° этап длился менее суток, при 3,5—3,7° — около 4 сут.
8 этап. Возникновение кардинальных вен и смешанного подкишечно-желточного и печеночно-желточного кровообращения. В первой серии между 49 и 52 сут. (195—207 градусо-дней) кровеносные сосуды эмбрионов обильно наполнились форменными элементами. Зародыши достигли длины 7,8—8,5 мм (рис. 41, B). Число сегментов в хвосте увеличилось до 35. Голова зародышей начала отделяться от желточного мешка. Расстояние между глазами и слуховыми капсулами сократилось до ширины 5 туловищных миотомов. Углубилась ротовая воронка. Появились зачатки нижних челюстей. Расширившаяся непарная плавниковая складка достигла 4—5-го туловищного сегментов. Кровью снабжалось 18—20 сегментов хвоста. Сформировались задние кардинальные вены, образовались кювьеровы протоки. Подкишечно-желточная вена опустилась ниже скопления жировых капель. На желточном мешке появилось 5—6 тонких капилляров.
При развитии в речных аппаратах сходное состояние кровеносной системы отмечено в конце 11 сут. — вначале 12 сут. (160— I65 градусо-дней), когда зародыши имели длину 7—7,3 мм.
Был проведен следующий опыт. Часть икры поместили в чашки Петри со слоем воды, едва закрывавшем икринки, и установили в речные аппараты. Оказалось, что в отсутствие проточности начало кровообращения задерживалось более чем на сутки. Сосуды развивались ненормально. По спинной аорте кровь текла только до середины туловнща, где сосуд выходил на желточный мешок, описывал короткую дугу и направлялся к сердцу. Позже кровь проникала в задний отдел туловища и в хвост, но эти сосуды развивались слабыми. Хвост формировался медленно, ненормально. Через некоторое время началась гибель икры. Перенос уцелевшнx икринок в проточную воду продлил жизнь зародышей, однако их развитие было уже нарушено и из таких икринок выходили уроды. Опыт показывает, что ослабление проточности при инкубации может являться одной из причин гибели икры и появления уродств.
У зародышей 1-й серии в возрасте 55 сут. (218 градусо-дней), а во 2-й — в 14 сут. (209 градусо-дней) начиналось печеночное кровообращение (по времени немного позже, чем у нерки и кижуча), появился ток крови в первой паре жаберных артерий, образо
вались первые сегментальные сосуды. В этом возрасте в глазах появился меланин. На рис. 41, Гизображен зародыш через 2 дня после появления в глазах меланина. Такие эмбрионы имели длину 9,5—9,9 мм и 77—78 миотомов. Их хвостовой отдсл составлял 30—30,5% всей длины. В почечных клубочках и мочеточнике появились кристаллы, что указывает на начало функционирования органа. Сегментальные сосуды образовались повсему туловищу и в первых сегментах хвоста. Сетью капилляров покрылась примерно половина поверхности желточного мешка.
В речных аппаратах в начале яигментации глаз зародыши имели длину 8,6—9,1 мм. У них сформировался 80—81 миотом, то есть сегментация закончилась. Кровью снабжались две пары жабер. Начала развиваться капиллярная сеть на дорзальной поверхности головы. При температуре 13,5—14° сердце сокращалось 50—51 раз в минуту.
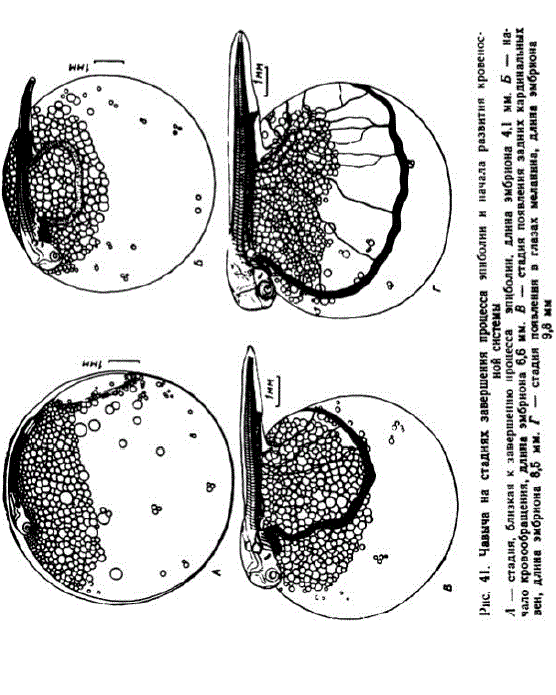
9 этап. Формирование печеночно-желточной системы кровообращения. При температуре около 4” течение крови из печени отмечено на 62—63-е сут. Эмбрионы достигли длины 10,3—11,8 мм и имели 78—80 миотомов (из них хвостовых 37—38), сегментация закончилась. Голова стала массивной. Слуховые капсулы увеличились, их основания расширились, образовались полукружные каналы и крупные отолиты. Кончик хорды начал изгибаться кверху и хвост утратил симметричность. Основания грудных плавников приняли наклонное положение. Наполнились кровью 1 или 2 пары жаберных артерий. Кровью не снабжалось только 6—8 последних хвостовых мнотомов. Капиллярами покрылся весь желточный мешок, исключая небольшой участок ниже головы. В течение этого этапа быстро увеличивалась длина хвоста.
У эмбрионов из речных аппаратов переход к печеночно-желточному кровообращению произошел на 18-е сут. (260 градусо-дней), когда они имели длину 10—11,5 мм (рис. 42, A). Кровью снабжались уже 3 пары жаберных дуг, наблюдался слабый ток крови в гиоидном сосуде. Появились капилляры на верхней поверхности головы. В большинстве хвостовых миотомов образовались сегментальные сосуды. Уплотнилась сеть капилляров на желточном мешке (на 1 мм2 его поверхности длина капилляров достигала примерно 4 мм).
10 этап. Формирование верхних и нижних конусов миотомов. Зачатки верхних и нижних конусов миотомов у зародышей первой серии появились на 68—70-е сут. (270—276 градусо-дней), эмбрионы имели длину 12—13 мм. Их голова стала выше, но продолжала оставаться относительно короткой. Хвостовой отдел составлял 3233% общей длины. Непарная плавниковая складка достигла наибольшей длины и заметно расширилась. Наметились зачатки пепарных плавников. Кровью снабжались три пары жабер. На дорзальной поверхности головы развилась густая сеть капилляров. Сосудами покрылась вся поверхность желточного мешка. Образовалась нижняя приносящая KCTO Hal Beha. Выявнлись первые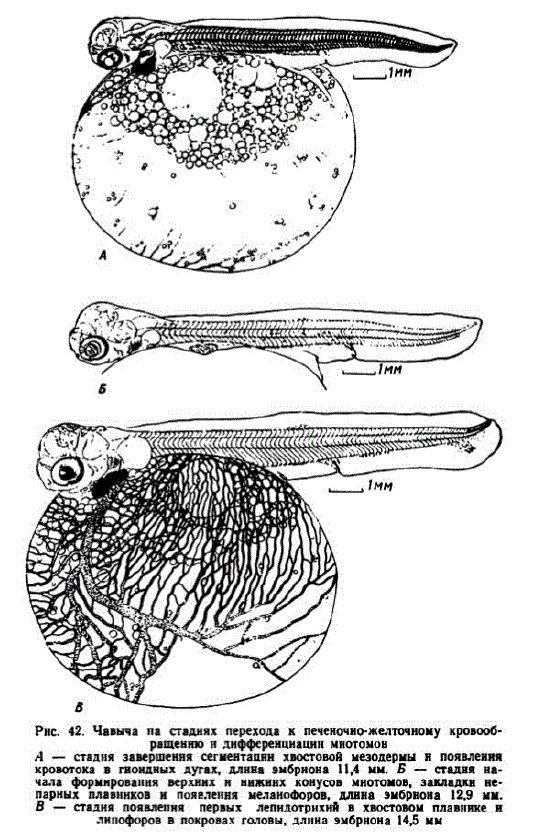
Pнс. 42. Чавыча па стаднях перехода к печеночно-желточному кровообращеннюнднфференцнацин мнотомон A — стадня завершення сегментацнн хвостовой мезодермы н появления кровотока в гноидных дутах, длнна эмбриона 11,4 мм. Б — стадня начала формировання верхних н нижннх конусов мнотомов, закладки непарных плавннков н появлення меланофоров, длнна эмбрнона 12,9 мм. B — стадня появлепня первых лепидотрихнй в хвостовом плавнике и лнпофоров в покровах головы, длина эмбрнона 14,5 мм
железки вылупления. На поверхности головы, вблизи крупных сосудов и на выстилке брюшной полости появились первые мсланофоры. На рис. 42, Бизображен эмбрион через несколько дней после начала этапа, развивавшийся в инкубаторе 75 сут.
У эмбрионов второй серии этап начался в возрастс 21 сут. (302 градусо-дня). Они имели длину около 13,5 мм и отличались более длинным хвостом (до 37% общей длины). Грудные плавники приобрели подвижность. В глазах появился гуанин, но еще отсутствовали меланофоры на голове и теле. Сегментальные сосуды сформировались во всех миотомах и в первых миотомах начали разветвляться.
Через 3 сут. такие зародыши достигли длины 14—15 мм (рис. 42, B; 344 градусо-дня). Их слуховые капсулы были максимально сближены с глазами, которые стали черными. Жаберные крышки прикрыли первую пару жабер. Нижние челюсти удлинились и, так же как жаберные крышки, приобрели подвижность. Верхние и нижние конусы развились на всех миотомах. Хвостовой отдел превысил 38% длины эмбрионов. Последнис хвостовые миотомы распадались и их число сократилось до 34—35. Сформировались три гипуралии. В хвостовом плавнике заложились первые лучи. Началось окостенение элементов плечевого пояса. Грудные плавники стали снабжаться кровью. Покрылась густой сетью капилляров верхняя поверхность головы. Кровеносные сосуды появились в основании лопасти хвостового плавника. На 1 мм2 поверхности желточного мешка длина капилляров превысила 7 мм. На данной стадии в покровах головы появляются малиново-красные липофоры (на рис. 42, Вони показаны черными мелкими точками).
Эмбрионы большинство времени оставались неподвижными, лишь изредка поворачивались с боку на бок. Периодически они совершали плавничками 10—15 взмахов в мин. По кишке время от времени проходили волны слабых перистальтических движений.
У зародышей, развивавшихся в инкубаторе, мускульные почки в основании спинного плавника появились на 76—79-е сут. (300— 310 градусо-дней). По сравнению с зародышами, развивавшимися в аппаратах с речной водой, они имели относитсльно укороченный хвост, длина которого составляла около 35% всей длины. Эти эмбрионы отличались также слабее развитой сосудистой сетью, а меланофоров имели больше. В возрасте 83 сут. (326 градусо-дней) зародыши достигли длины 13,5—14,7 мм. У них заложились брюшные плавники. Грудные плавникн приобрели подвижность. В просвете кишки был виден желто-зеленоватый пигмент. Повысилась
BH3 KOOTH MIKI.TK.
11 этап. Развитне подвижности челюстей, жаберных крышек, завершение инкубации. В первой серии у разных особей этап начинался на 90—95-с сут., а во второй — на 26-е сут. (соответственпо 347—364 и 372 градусо-дня). Первые эмбрионы имели длину 14,8—16,3 мм. У них начал увеличиваться относительный размер головы. Жаберные крышки почти прикрыли вторую пару жабер
Появились зачатки жаберных лепестков. В обонятельных отверстиях наметилось образование персмычек. Хвостовой отдел составлял 36% всей длины эмбрионов. Образовались гипуралии и стало больше лучей хвостового плавника. В анальном и спинном планниках наметилась дифференцировка опорных элементов. Заложились брюшные плавники. Сегментальные сосуды развились по всему телу. В головной чаституловища появились капилляры над спинным мозгом и у поверхности тела. Длина капилляров, располагающихся на 1 мм2 поверхности желточного мешка, достигла 6 мм. Сосудистая сеть у этих особей была слабее, нежели у одноразмерных эмбрионов, развивавшихся в речных аппаратах (см. рис. 43, А, Б). Хвост последних был длиннее, свыше 38% всей длины, а его лопасть развита лучше. Следует отметить произошедшее усиление пигментации, причем меланофоров было больше на головс и в хвосте, нежели в туловищной области. Интересно, что узародышей из инкубатора только через 70 градусо-дней после появления меланофоров в покровах головы были обнаружены первые липофоры. У тех же зародышей, которые развивались в речных аппаратах, первыми обнаруживались липофоры и примерно через 40 градусо-дней после этого, в возрасте 26 сут. (рис. 43, A), выявились меланофоры. Так как материал сравниваемых серий генетически однородный, отмеченные различия свидетельствуют о зависимости процесса пигментации от условий инкубации икры.
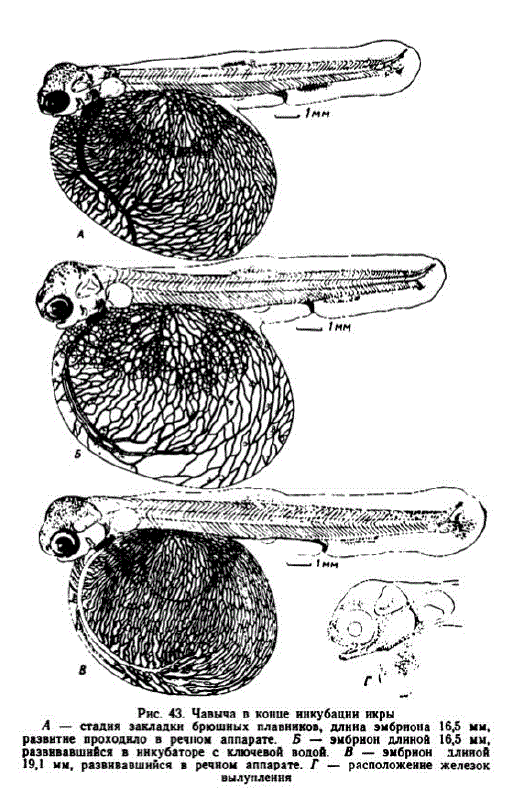
На 31-е сутки эмбрионы в речных agai JLOCTHTIH IIHHLN около 19 мм (рис. 43, B; 440 градусо-дней). Их жаберные крышки закрывали 3 пары жабер. В жаберные лепестки начали врастать капилляры. В этом возрасте хвостовой отдел составлял до 40% длины эмбрионов. В хвостовом плавнике образовалось 12—14 лучей; самые крупные из них имели по 2 членика. Между лучами развилась густая сеть капилляров. В хвосте начали формироваться позвонки. Продолжалась дифференцировка анального и спинного плавников. Появились признаки редукции непарной плавниковой складки. Сосудистая сеть на желточном мешке достигла максимального развития. На 1 мм2 его поверхности длина капилляров превышала 9 мм.
Находясь под оболочкой, такие эмбрионы почти беспрерывно работали грудными плавниками, совершая при 14,5° 180—200 взмахов в минуту. Сердце сокращалось 90—95 раз в минуту. Пернодически при возбуждении они производили волнообразные трспещущие движения хвостом. Извлеченные из икринок, такие зародыши моглн проплыть небольшое расстояние.
К 38 суткам (527 градусо-дней) эмбрионы этой серни достигли длины 20—21 мм. Голова составляла 15% длины тела. Нижние челюсти разрослись до вертикали, опущенной от линии переднего края глаз. В ротовой полости развился velum. Эмбрионы ритмично двигали челюстями, жаберными крышками и жабрами. В зачатках псевдобранхий развилась сеть капилляров. Разросшиеся перемычки почти полностью разделили обонятсльные отверстия.
В хвосте образовалось 15—17 лепидотрихий; наиболее крупные из них имели по 3—4 членика. Отчетливо выявились контуры непарных плавников. Поверхность головы зародышей стала темной. Меланофоры распространились повсему телу, но их было заметно меньше в передней третитуловища. Масса пигментных клеток покрыла промежутки между лучами хвостового плавника. Появились хроматофоры в лопасти анального и в основании жирового плавников. Четко выявились многочисленные железки вылупления (рис. 43, Г). По сравнению с неркой, у чавычи железок больше на рыле, нижних челюстях и грудных плавниках, но меньше нажаберных крышках.
При хорошем водоснабжении аппаратов такис эмбрионы под оболочкой двигаются редко. Грудными плавниками при 13—13,5° они совершали до 190—200 взмахов в минуту; сердце сокращалось 89—91 раз в 1 минуту. Перед вылуплением эмбрионы возбуждались, начинали вращаться подоболочкой. Интенсивность их дыхаH H H IIOBECHIACh.
12 этап. Пассивное состояние свободных зародышей. Вылупление зародышей в речных аппаратах в 1956 г. началось на 39-е сут. (537 градусо-дней), массовым стало на 41-е сут. (563 градусодня) и закончилось через 3 сут. На следующий год вылупление началось на 34-е сут. (470 градусо-дней) и было массовым на 36-е сут. Для чавычихарактерен дружный выход из икры. В инкубаторе в 1956 г. групповое вылупление проходило в возрасте 145— 150 сут. (500—513 градусо-дней) и закончилось в возрасте около 160 сут. (540 градусо-дней). В 1957 г. отмечено более раннее вылупление, начавшееся на 101-е сут. (397 градусо-дней). На Ушковском заводе в 1934—1939 гг. массовое вылупление чавычи проходило в среднем через 95 сут. после оплодотворения (около 576 градусо-дней).
В 1957 г. при групповом вылуплении в инкубаторе зародыши имели длину 18—20,3 мм (в среднем 19,4 мм) и всcили 212—214мг (в среднем 228,4 мг). Вес желтка составлял 165—189 мг, в среднем 180 мг, а средний вес тела эмбриона — 49 мг, что немного более 21% общего веса. В речных аппаратах зародыши вылуплялись при длине 19—21 мм (в среднем 19,9 мм) и весе 216—251 мг (в среднем 233 мг). Вестела эмбрионов колебался от 42 до 74 мг и в среднем составлял около 60 мг (немного больше 25% общего веса). За период инкубации в условиях повышенной температуры вес желтка уменьшался на 12,5%.
Вид зародыша в период массового вылупления иллюстрирует рис. 44, А. В 1956 г. икра была крупной и зародыши при вылуплении имели длину 22—23 мм. Желточный мешок зародыша яйцевидный. Большинство жировых капель слились, образовалась одна огромная капля. На рисунке показано их расположение у зародыша, лежавшего на боку. Когда зародыши держатся спинкой кверху, жировые капли занимают дорзальное положенис.
Во время вылупления насчитывалось 72—75 миотомов, хвост
имел 29—31 миотом и составлял около 41% всей длины. Рот зародышсй полунижний. Глаза крупные, неподвижные. Разросшиеся кожные выросты жаберных крышек не покрывали лишь кончики некоторых жаберных лепестков. В псевдобранхиях развились круп"IEC JOACTI: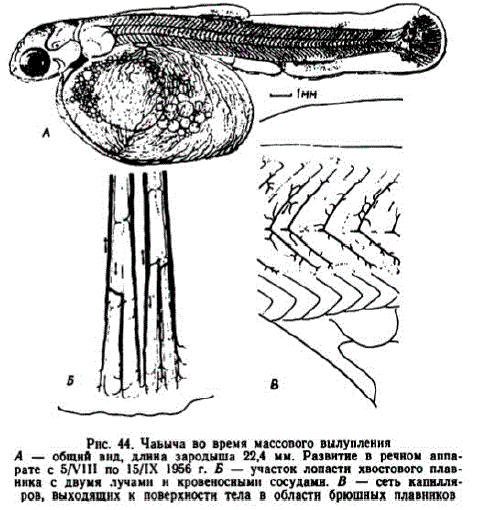
Рис. 44. Чаьыча во время массопого вылупления A — общнй внд, длина зародыша 22,4 мм. Развитне в речном аппарате с 5/VIII по 15/IX 1956 г. Б — участок лопастн хвостового плавHHKH двумя лучами H кровеносными сосудамн. B -- EPT KaILLIFров, выходящих к поверхности тела в областн брюшных плавников
В непарных плавниках продолжалась дифференцировка лучей, появились меланофоры и липофоры, четко выявились их контуры; oбозначился контур и жирового плавничка. В хвостовом плавникс насчитывалось до 20—22 лучей (при раннем вылуплении имеется только 13-15 лучей). Вершины брюшных плавников почти достигали края преанальной плавниковой складки. Появились зачатки лучей в грудных плавниках. Тело вылупившихся зародышей пигментировано интенсивнее, чем у кеты, горбуши и нерки.
После вылуплення тонкие капилляры желточного мешка начинают запустевать. Эта утрата частично компенсируется развигием капилляров между лучами хвостового плавника (рис. 44, Б) и
вблизи поверхности тела (рис. 44, В). Существенно возрастает роль жабсрного дыхания.
На осторожнос прикосновение зародыши не реагировали, также как и на слабое освещение. Яркий свет возбуждал зародышей, но к слабому свету они относились безразлично. Вылупившиеся заpoliIII IenOLBIKHO TeKaT Ha U.LIOMI MeCTU, энергично помахивают трудными плавниками. При 14—15° сердце сокращается 92—95 раз в минуту. Поднятыс струей воды, зародыши короткое время могут активно удерживаться в толще воды. После выхода из икринок потребление зародышами кислорода резко возрастает (см. табл. 13).
13 этап. Формирование непарных, брюшных плавников и плавательного пузыря. Через некоторое время, аналогично другим видам, поведение зародышей изменяется. В 1957 г. в речных аппаратах это было заметно на 46-е сут. (617 градусо-дней), когда зародыши имели длину 22—24 мм (средняя 22,9 мм). Они начали перемещаться навстречу течению, положительно реагировали наприкосновение посторонних предметов, проявляли сильное беспокойство на освещенных участках и прятались в темноту. При температуре 2,5—2,6° аналогичные изменения происходили приблизительно через 15 дней после вылупления, причем в этих условиях смена поведения проявлялась менее четко.
В речных аппаратах через 18 сут. после вылупления зародыши достигали длины 28,2—29,7 мм (за это время температура снижалась с 12 до 9,5°). На их теле появились пятна (рис. 45, А; развитие с 5/VIII по 3/X 1956 г., 755 градусо-дней). Непарная плавниковая складка сильно уменьшилась. Лучи спинного плавника стали расчленяться. Хвост приобрел симметричную форму. Появились так называемые редуцированные хвостовые лучи. Образовались лучив грудных и брюшных плавниках. Почти третью часть желточного мешка закрыли разрастающиеся миотомы. В течение этапа наблюдался быстрый рост зачатка плавательного пузыря. В аппаратах чавыча периодически поднималась в толщу воды, но удерживалась на плаву короткое время. В условиях низкой температуры пятна на боках тела чавычи появились в возрасте 185 суток (610 градусо-дней).
Смотрите также
Бассейновый метод выращивания лососевых рыб