
ИЗМЕНЧИВОСТЬ МОРСКОГО ПЕРИОДА ЖИЗНИ АТЛАНТИЧЕСКОГО ЛОСОСЯ НА СЕВЕРО-ВОСТОКЕ АРЕАЛА
Биологические группы. Созревание производителей атлантического
лосося по «озимому» типу рассматривается как адаптация к воспроизводству
в протяженных речных системах (Берг, 1935; Шмидт, 1947). Действительно,
в крупных лососевых реках доля «озимых» рыб велика и может
достигать 100 %. Однако есть относительно короткие реки (Кола,
Йоканьга, Варзуга, Выг, Нива, Онега и др.), в которые заходит, или в свое
время заходило, много «озимых» рыб. В этом случае вряд ли можно рассматривать
созревание рыб по «озимому» типу как приспособление к
преодолению только пресноводного участка миграционного пути, поскольку
в перечисленных реках нижняя граница НВУ располагается в нескольких
десятках, а то и в километрах от моря. С другой стороны, основные
морские районы нагула и зимовки указанных популяций находятся в
южной части Норвежского моря и отстоят от мест размножения на многие
сотни километров, преодоление которых требует существенных затрат
времени и энергии. Большую роль протяженных морских миграций в
формировании сезонных рас проходных тихоокеанских лососей, в частности
амурской кеты, отмечал И.Б. Бирман (1981). Протяженность морского
отрезка миграционного пути для восточно-европейских популяций
атлантического лосося изменяется примерно от 0,8 (реки западного Мур-
мана) до 3,2 тыс. км (реки на востоке ареала). Затраты времени лососей
при совершении нерестовых миграций, наряду с затратами энергии, на
наш взгляд, имеют существенное значение в формировании их межпопу-
ляционной дифференциации. Одно дело, когда небольшие лососевые реки
находятся в пределах морских районов нагула, и совершенно другое, когда
НВУ в верховьях реки отделены от морских районов нагула и зимовки
протяженным магистральным руслом (реки Висла, Рейн и др.) или же в
дополнение к этому не менее протяженным пространством океана (реки
Печора, Северная Двина, Мезень). В первом случае для достижения нерестилищ
потребуется минимальное время, а в остальных затраты времени
будут исчисляться многими десятками дней.
Среднюю скорость лососей, совершающих морскую преднерестовую
миграцию, можно принять равной 30 км/сут (Шмидт, 1947; Азбелев, 1960;
Бакштанский и др., 1991; Потуткин, 2004), а скорость в реке - 33 км/сут
(Новиков, 1956; Летовальцева, 1967). Тогда, например, для преодоления
расстояния от Фарерских островов до устья р. Печоры рыбам потребуется
3 мес. и дополнительно 1,5-2,0 мес. на прохождение пресноводного учас-
тка миграционного пущ. Если учесть, что в реке половые продукты зашедших
из моря рыб созревают в течение 2 мес, то суммарный бюджет времени
на нерестовую миграцию и формирование половых продуктов для печорского
лосося составляет 7 мес. Период же с благоприятными условиями
для миграции и нереста на крайнем востоке ареала не превышает 5 мес.
(май-сентябрь). Как видим, для реализации бюджета времени на воспроизводство
в нашем примере потребуется два вегетационных сезона, что в
состоянии осуществить только рыбы, созревающие по «озимому» типу.
Косвенным подтверждением значимости морских условий жизни в
формировании биологических групп в популяциях атлантического лосося
может служить отсутствие «озимых» рыб на Североамериканском континенте.
Северо-американский лосось нагуливается у западных берегов
Гренландии до 65°30' с. ш., а зимует в районе между 42° и 55° с. ш. на
удалении примерно 250-600 км от Ньюфаундленда (Reddin, Shearer, 1987;
Reddin, 1988; Reddin, Friedland, 1993), т. е. в относительной близости к нерестовым
рекам. В этих условиях бюджет времени на воспроизводство
сокращается за счет уменьшения его «морской» составляющей и может
быть реализован рыбами летней биологической группы. Протяженность
же пресноводного участка миграционного пути для североамериканских
популяций атлантического лосося не превышает 400 км (Schaffer, Elson,
1975). Распространение лосося в наиболее протяженной р. Сент Джон
(цит. по: Джефард, 1998) до ее зарегулирования ограничивалось порогами,
расположенными примерно в 400-500 км от устья.
Таким образом, созревание атлантического лосося на северо-востоке
ареала по «озимому» типу может рассматриваться прежде всего как приспособление
к большим затратам времени на миграцию из морских районов
нагула к нерестилищам в условиях относительно узкого «миграционного
окна». Вместе с тем рыбы осенней биологической группы совмещают
морскую преднерестовую миграцию с нагулом, и их последние морские
приросты длины и массы значительно превышают таковые «яровых
» рыб, приближаясь по величине к годовым (см. гл. 6). Данное обстоятельство,
а также более высокая жирность рыб осенней биологической
группы (Тилик, 1935) позволяют рассматривать созревание лососей
по «озимому» типу как адаптацию двойного назначения: к затратам времени
и энергии на миграцию к нерестилищам и воспроизводство.
Размеры и возраст. На примере атлантического лосося из рек Канады
(летняя биологическая группа) показано (Schaffer, Elson, 1975), что средняя
масса рыб в популяциях положительно коррелирует с длиной нерестовых
рек и носит адаптивный характер к уровню энергетических затрат
на преодоление пресноводного участка миграционного пути. Об уровне
энергетических затрат лососем на преодоление пресноводного участка
миграционного пути можно судить по потерям массы нерестовых мигрантов
в бассейне Печоры (Мартынов, 1983). Преодоление анадромны-
ми мигрантами щугорского лосося 1000-километрового отрезка магист-
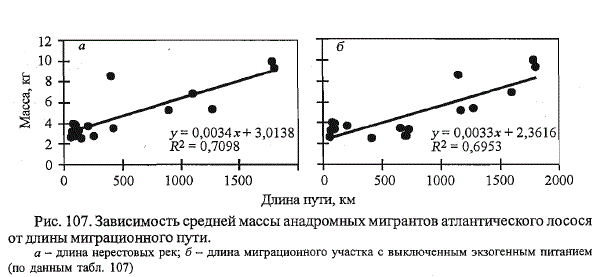
Рис. 107. Зависимость средней массы анадромных мигрантов атлантического лосося
от длины миграционного пути.
а - длина нерестовых рек; б - длина миграционного участка с выключенным экзогенным питанием
(по данным табл. 107)
Fig. 107. The mean weight of angled fish plotted against migratory way length.
a - length of rivers; 6 - length of migratory way without food. Data from table 107
ральной реки в течение месяца сопровождается потерей 23 % массы тела,
что составляет более половины суммарных потерь за время пребывания
в реке и нерест (см. гл. 4).
Средняя масса производителей в популяциях лосося на Севере России,
так же как и в популяциях атлантического лосося Канады, положительно
коррелирует с длиной нерестовых рек (рис. 107, а). Из общей картины
выделяется лосось р. Онеги, отличающийся высокими размерно-
весовыми характеристиками при относительно небольшой длине нерестовой
реки. Если учесть, что при заходе в Белое море нерестовые мигранты
лосося перестают питаться (см. гл. 6), и суммировать с длинами нерестовых
рек беломорского лосося протяженность миграции рыб в Белом
море, то данный феномен пропадает (см. рис. 107, б). Таким образом,
высокие размерно-весовые показатели онежского лосося можно рассматривать
в контексте повышенных затрат энергии рыбами на прохождение
протяженного беломорского участка миграционного пути, к тому же преодолеваемого
ими против течений, существующих в Белом море.
Продолжительность морского периода жизни лосося положительно
коррелирует с размерами нерестовых мигрантов (см. гл. 3), и ее изменчивость
следует рассматривать в контексте адаптированности рыб в популяциях
к уровню затрат энергии на миграцию к нерестилищам и воспроизводство.
Взаимоотношения полов. Таким образом, увеличение затрат энергии
на воспроизводство может компенсироваться при переходе лососей
на более длительный морской нагул и, частично, созревание по «озимому
» типу, что приводит к соответствующим изменениям в структуре популяций.
Однако самки и самцы атлантического лосося на Севере России
по мере продвижения на восток и увеличении протяженности миграционного
пути по-разному реагируют на меняющиеся условия обитания
(см. гл. 3 278 и табл. 107). На наш взгляд, в основе данного явления лежат
Таблица 107. Протяженность миграционного пути и средняя масса производителей
атлантического лосося на Севере России
Table. 107. Extent of migration way and average weight of Atlantic salmon in the North of
Russia
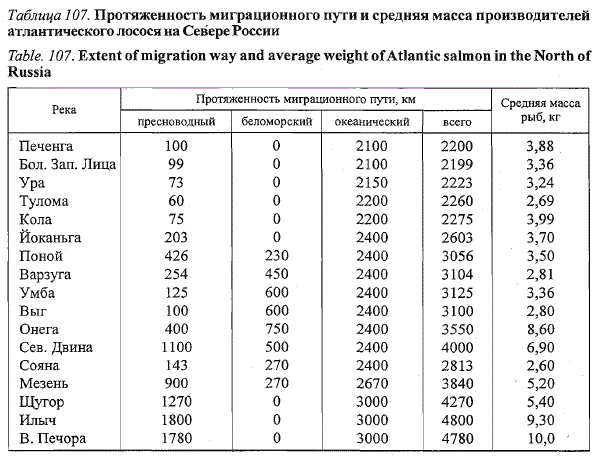
Примечание. Таблица составлена по данным, представленным в гл. 3.
существенные различия между рыбами разного пола в энергетических
затратах на формирование половых продуктов. Как показано Джонсоном
с соавторами (Jonsson et al., 1991), доля гонад от общей массы тела у половозрелых
лососей, выраженная в энергетическом эквиваленте (кДж), составляет
у самок 27,8 %, а у самцов — 4,5 %, т. е. энергетические затраты
самок на формирование половых продуктов в 6,2 раза выше, чем у самцов.
При одинаковых размерах с самками и соответственно сопоставимом
уровне энергозапасов самцы располагают большими возможностями
на энергозатраты, не связанные с генеративной функцией, и при выключенном
экзогенном питании в состоянии преодолевать без ущерба
формированию половых продуктов значительно большие расстояния. Поэтому
с удлинением миграционного пути к нерестилищам самки начинают
испытывать недостаток энергетических запасов при меньших размерах,
чем самцы.
В «яровых» популяциях лосося, воспроизводящихся в относительно
небольших речных системах недалеко от основных морских районов нагула
(реки западного Мурмана), возникающее противоречие разрешается
в результате перехода большинства самок на двухлетний морской нагул, в
то время как самцы созревают в основном как тинда. При возрастании
279
энергетических затрат, не связанных с формированием половых продуктов,
до уровня, близкого к верхней границе адаптационных возможностей
«яровых» самок с двухлетним периодом морского нагула, возникает
предпосылка к их созреванию после трех лет морского нагула. Однако
такая адаптация на первых порах, по-видимому, является избыточной,
ведет к снижению темпа воспроизводства и слабо поддерживается отбором.
В условиях возникновения дефицита времени самки начинают переходить
к созреванию после одного года морского нагула по «озимому»
типу. В популяциях лосося появляется заледка (реки восточного Мурма-
на), доля которой по мере продвижения к востоку ареала возрастает и
достигает максимального значения в популяции лосося р. Варзуги, в то
время как большинство самцов продолжает созревать по «яровому» типу
как тинда. В результате в бассейне р. Варзуги, на достаточно большом
удалении от основных морских районов нагула, сформировалась своеобразная
популяция лосося, почти полностью представленная рыбами с
одним годом морского нагула, в которой наиболее ярко проявляется половой
диморфизм по типу созревания.
При воспроизводстве на еще большем удалении от морских районов
нагула самки лосося вновь переходят к созреванию после двух лет морского
нагула, но уже как «озимые» рыбы. Вслед за ними созревать по «озимому
» типу после одного и двух лет морского нагула начинают самцы.
И наконец, при воспроизводстве на самых удаленных от морских районов
нагула нерестилищах, расположенных в бассейне Верхней Печоры,
все самки и самцы лосося созревают по «озимому» типу после двух и
более лет морского нагула.
В популяциях лосося на Севере России анадромные мигранты, особенно
самцы, представлены особями, сильно различающимися по размерам.
Не исключено, что подразделение самцов в популяциях лосося на
мелких и крупных может быть связано с особенностями влияния размерно-
весовых параметров производителей на жизнеспособность потомства.
Как показано на примере горбуши, кеты и нерки р. Большой, относительно
низкими показателями смертности на поздних этапах эмбрионального
развития характеризовалось потомство от мелких (младших) и крупных
(старших) самцов, в то время как средневозрастные самцы давали
потомство с относительно высокой смертностью (Чебанов, 1984). Более
высокая выживаемость потомства была отмечена для балтийского лосося
(Казаков, 1979) при оплодотворении икры спермой младшевозраст-
ных самцов. Относительно высокая выживаемость потомства мелких и
крупных самцов в популяциях проходных лососей может быть одним из
механизмов повышения внутрипопуляционной гетерогенности размерно-
весовых характеристик рыб. В частности, бимодальное распределение
самцов по морскому возрасту в популяциях атлантического лосося
западного Мурмана (см. рис. 60), скорее всего, обусловлено более высоким
выживанием потомства мелких (Р+1+) и крупных (Р+3+) рыб.
Различия в размерно-возрастной структуре, типе полового созревания
(биологические группы, карликовость), характере роста между самками
и самцами в той или иной степени проявляются во всех исследованных
популяциях лосося на Севере России. Они воспроизводятся в
поколениях и устойчивы во времени. Наличие комплекса биологических
особенностей у рыб разного пола и специфика их географической изменчивости
дают основание рассматривать самцов и самок атлантического
лосося аналогично самцам и самкам проходных дальневосточных лососей
(Алтухов и др., 1997) как относительно самостоятельные взаимообусловленные
внутривидовые адаптивные системы.
Смотрите также