ИЗМЕНЧИВОСТЬ РЕЧНОГО ПЕРИОДА ЖИЗНИ АТЛАНТИЧЕСКОГО ЛОСОСЯ НА СЕВЕРО-ВОСТОКЕ АРЕАЛА
Размеры и возраст. Факторы, определяющие внутрипопуляционную
изменчивость размерно-возрастной структуры покатников атлантического
лосося, могут быть абиотической и биотической природы. Взаимодействуя,
они определяют особенности размерно-возрастной структуры
покатников в различных реках.
Длительность речного периода жизни лосося в реках севера России
отрицательно коррелирует с теплосодержанием воды на НВУ «родных»
рек (табл. 104, рис. 104). В основе положительной зависимости между
средней продолжительностью речного периода жизни атлантического
лосося и широтой местности, видимо, также лежит отрицательная связь
между температурой, понижающейся с увеличением широты, и длительностью
пребывания молоди в реке. Внутрипопуляционная изменчивость
возрастной структуры молоди в речной период жизни может быть свя-
Таблица 104. Средняя продолжительность речного периода жизни атлантического лосося
и сумма градусо-дней воды в реках Севера России
Table 104. Average river age of Atlantic salmon and the sum of water temperature-days in the
rivers of the North of Russia
Примечание. Сумму градусо-дней определяли как среднее из годовых значений по гидрометеопо-
стам, расположенным в бассейне соответствующей реки в пределах НВУ атлантического лосося (см.
Приложение, табл. 1).
зана со степенью гетерогенности температурного режима на микростациях,
занимаемых пестрятками. В вегетационный период определенная
мозаичность температурного режима на них формируется под влиянием
инсоляции и поступления охлажденных грунтовых вод. На НВУ
в пределах порогов в прибрежной полосе, где на малых глубинах скорость
течения замедлена, температура воды за счет инсоляции обычно
на 3-4 °С выше, чем в центральной части порога. Небольшой размах
возрастного ряда покатников лосося наблюдается в крупных реках Центральной
геоморфологической провинции, характеризующейся широким
развитием карстовых явлений и сглаженным температурным режимом.
Увеличение размаха возрастного ряда покатников в малых ре-
Рис. 104. Зависимость средней продолжительности
речного периода жизни атлантического
лосося на Севере России от теплосодержания
воды в нерестовых реках (по данным
табл. 104)

Fig. 104. Dependence of average duration
of the river life period of Atlantic salmon in the
North of Russia on water temperature in
spawning rivers. Data from Table 104
ках Мурмана может быть связано не только с абиотическими, но и биотическими
условиями.
На Севере России средний показатель смертности в период от ската
молоди лосося в море и до возврата взрослых рыб на нерест в реки достигает
95 % (Бакштанский и др., 1976). Очевидно, что большая доля естественной
смертности в этот период приходится на покатную миграцию
и ранний период морской жизни, когда относительно небольшие размеры
делают молодь более доступной для хищников. Основным туводным
хищником, питающимся молодью лосося в период покатной миграции,
является щука, которая широко распространена на Севере России и встречается
во всех лососевых реках (см. гл. 5). К дополнительным хищникам
со сходной экологией питания можно отнести крупную речную кумжу (р.
Йоканьга) и арктического гольца (р. Варзино). Эти три вида могут быть
существенным фактором элиминации покатной молоди лосося. Налим,
который также широко распространен в лососевых реках, потребляет молодь
лосося на стадии пестрятки. Его питание покатной молодью не отмечено.
В р. Умба щука выедает до 35 % покатной молоди лосося (Кузьмин и
др., 1985). Как показано для р. Кереть, на питание выпущенной в реку
заводской молодью лосося в основном переходят средние по размерам
щуки массой от 1-2 кг (Костылев, Ермолаев, 1982а). Ими выедается 25-
30 % заводской молоди, выпущенной в реку (Бугаев и др., 1986). В малой
беломорской р. Порье щукой истребляется до 1/3 диких покатников (Смирнов
и др., 1977).
В целом ряде исследований, обзор которых приводится в монографии
Ю.Ю. Дгебуадзе (2001), показано повышение элективности водных хищников
по мере увеличения (до определенного уровня) размеров жертв.
Однако следует подчеркнуть, что в большинстве из рассмотренных работ
характер элективности хищников устанавливался по отношению к беспозвоночным.
B.C. Ивлевым (1955) в эксперименте показано увеличение
элективности щуки в отношении среднеразмерных особей воблы.
Предпочтение щукой более мелких рыб выявлено в экспериментальных
условиях при использовании в качестве жертв молоди красноперки (Hart,
Hamrin, 1988). При сопоставлении размерных характеристик заводской
молоди балтийского лосося, помеченной перед выпуском в реку, с соответствующими
показателями меченой молоди, обнаруженной в желудках
щук, установлено, что в естественных условиях хищником поедается более
мелкая молодь (Larsson, 1977). На примере карповых рыб Саратовского
водохранилища (чехонь, синец, уклея, плотва, лещ, густера) показано,
что хищниками (щука, сом, судак, жерех, окунь, берш, налим) в наибольшей
степени выедаются мелкие особи младших возрастных групп. С увеличением
размеров жертв степень их выедания хищниками снижалась и
при достижении длины 30 см достигала нулевых значений (Ермолин,
1979).
В природной среде взаимоотношения между покатной молодью лосося
и основным туводным хищником щукой носят сложный коадаптив-
ный характер (Бакштанский, Нестеров, 1976; Бакштанский и др., 1980а,
1990; Bakshtansky, Nesterov, 1974; Bakshtansky et al., 1977). Показано, что
поведенческие реакции потенциальной жертвы на меняющиеся абиотические
условия среды носят выраженный адаптивный характер и в конечном
счете направлены на избегание хищника. С другой стороны, в
период катадромной миграции молоди лосося хищник способен менять
свой обычный ритм суточной активности, согласуя его с суточной ритмикой
миграции жертвы. При этом существенным обстоятельством является
взаимодействие хищника не с одиночными особями жертвы, а с
их стайными образованиями (Бакштанский и др., 1978).
Выедая более мелких рыб, хищники, несомненно, способствуют формированию
особенностей размерной и возрастной структуры покатной
молоди в популяциях лосося. Роль данного фактора может быть выявлена
при сопоставлении размерно-возрастной изменчивости покатников в
реках с высокой и низкой численностью засадных хищников. В качестве
выраженных крайних вариантов можно рассмотреть размерно-возрастную
структуру покатников лосося озерно-речной системы Иоканьги, где
засадные хищники представлены многочисленными щукой и кумжей, и
Печоры, в безозерных лососевых притоках которой обитает малочисленная
щука. Особенности размерно-весовой изменчивости йоканьгских
покатников состоят почти в полном совпадении размаха колебаний и
небольших различиях средних показателей длины и массы тела у рыб всех
возрастных групп (рис. 105, а; см. табл. 78). Средние показатели размеров
молоди печорского лосося на стадии покатника существенно различаются
не только в крайних, но и смежных возрастных группах, а пределы
колебаний размеров с увеличением возрастной категории закономерно
смещаются вправо (см. рис. 105, б, табл. 84). Изменчивость размерно-
возрастных параметров покатной молоди лосося других исследованных
нами рек занимает промежуточное положение. У лосося рек Мурмана по
своему характеру она близка к «йоканьгскому», а в ряде беломорских рек
(Варзуга, Чаваньга, Стрельна) - к «печорскому» типам.
Если принять за обобщенный показатель размерно-возрастной изменчивости
покатников разницу средних длин рыб в крайних возрастных
группах и соотнести их с суммарной численностью засадных хищников
на НВУ лосося (табл. 105), то между данными параметрами выявляется
обратная зависимость (рис. 106): чем выше численность хищников, тем
меньше фенотипические различия между рыбами разных возрастных
групп. При этом размерное сходство между ними достигается через возрастные
различия в темпе роста, когда младшевозрастные покатники зачастую
растут быстрее старшевозрастных в несколько раз (см. раздел 4.3).
Выедая мелких покатников, селективные хищники в конечном счете
осуществляют отбор по темпу роста. У крупных рыб, которые могут быть Рис.
Рис.
105. Гистограммы распределений по длине разновозрастных покатников
атлантического лосося рек Йоканьги (а) и Печоры (б). Результаты обратного расчисления
по чешуе производителей
Fig. 105. Length-frequency distributions of Atlantic salmon smolts in the Iokanga (a) and
Pechora (6) Rives by age. Results of beak calkulations on scales of adult fisch
представлены, с одной стороны, младшевозрастными быстро растущими
особями, а с другой - тугорослыми старшевозрастными, появляется
больше шансов выжить. Отбор рыб с более высоким темпом роста будет
происходить во всех возрастных группах, однако младшевозрастные, а
следовательно, более мелкие покатники будут испытывать больший пресс
хищников, чем старшевозрастные. Таким образом, положительный отбор
по темпу роста окажется более сильным в младшевозрастных группах
покатников. При длительном воздействии отбора должно произойти
сближение средних показателей длины рыб в младших и старших возрастных
группах, причем степень конвергенции будет зависеть от силы давления
отбора, которая пропорциональна численности хищников.
Кроме того, в результате отбора младшевозрастных рыб с высоким
темпом роста и старшевозрастньгх тугорослых рыб происходит растяжение
возрастных рядов покатников, что мы и наблюдаем в популяциях атлантического
лосося, воспроизводящихся в насыщенных хищниками от-
Таблица 105. Возрастная изменчивость длины тела покатников атлантического лосося
в реках на Севере России (наши данные)
Table 105. Age variability of length of Atlantic salmon smolts in the rivers of the North of
Russia (our data)
Примечание. В скобках указано количество экземпляров. Суммарная численность хищников приводится
на основе табл. 42 Приложения; данные по р. Стрельна - Долотов, 1997, по р. Йоканьга -
Зубченко и др., 1991.
носительно небольших озерно-речных системах Мурмана, где покатники
представлены рыбами до шести возрастных групп (от 2+ до 7+). В крупных
безозерных лососевых реках с низкой плотностью хищников покат-
ная молодь лосося представлена 3-4 возрастными категориями.
Высказаное П.И. Новиковым (1953) положение о том, что молодь лосося
скатывается в море по достижении определенного размера и в значительной
мере независимо от возраста было основано на том факте, что
быстро растущая молодь мигрирует в море в более раннем возрасте, чем
тугорослая, но при близких с ней средних показателях длины. Данное
положение получило название «концепции предопределенности размеров
покатника» (Дирин, 1980). Наличие популяций лосося, в которых на-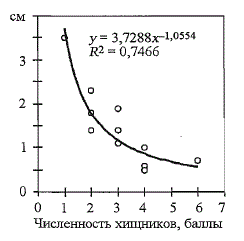
Рис. 106. Связь между размерно-возрастной изменчивостью
покатников атлантического лосося
(разница средних показателей длин у покатников в
крайних возрастных классах) и численностью тувод-
ных хищников (по данным табл. 105)
Fig. 106. The difference of average length in minimal
and maximal age groups of fish (on ordinate) plotted
against predator number (on abscissa) for salmon rivers
in the North of Russia. Data from Table 105 1 - -
блюдаются существенные различия средних
показателей длины и массы разновоз-
с- « Численность хищников, баллы
растных покатников, а также большой раз-
мах колебаний размеров рыб в пределах
возрастных групп послужили основанием для критики представления о
предопределенности размеров смолта (Азбелев, 1960; Дирин, 1980). Вместе
с тем на уровне вида размеры, при которых наступает смолтифика-
ция, несомненно, наследственно детерминированы, и межпопуляцион-
ные различия длины и массы покатников демонстрируют изменчивость
генома вида в разных условиях ареала. Так, средняя масса покатников
лосося на Севере России различается почти в 4 раза: от 10 г в р. Варзуге
до 39,7 г в р. Йоканьге. Причем в р. Варзуге разновозрастные покатники
существенно различаются по средним показателям длины и массы, демонстрируя
положительную связь между возрастом и размерами смолта
(см. табл. 82), а в р. Йоканьге такие различия практически отсутствуют,
что может быть истолковано в пользу представления о предопределенности
размеров покатника (см. табл. 78). На наш взгляд, степень выраженности
феномена предопределенности размеров покатника в разных
популяциях атлантического лосося во многом может быть связана со степенью
воздействия хищников.
Анализ роста молоди лосося (см. гл. 4) свидетельствуют о его более
высоком темпе в баренцевоморских реках. Самый высокий темп роста
молоди лосося в р. Йоканьге, характеризующейся относительно низким
теплосодержанием воды, на наш взгляд, может быть в определенной степени
связан с воздействием хищников. Наиболее низкий темп роста молоди
лосося в р. Варзуге с относительно высоким теплосодержанием воды,
по мнению Э.Л. Бакштанского и В.Д. Нестерова, высказанному ими при
обсуждении данной работы, может быть результатом популяционной
адаптации к особенностям условий воспроизводства в этой реке, характеризующейся
высокой долей в составе НВУ нерестовых площадей. По
данным Р.В. Казакова с соавт. (1992), доля нерестовых участков лосося в
общей площади НВУ р. Варзуги составляет 23 %, что в 2,3 раза превышает
соответствующий показатель для р. Онеги (Кулида, 1982) и в 3,3 раза—
для североуральской р. Щугор (Мартынов, 1979, 1983). При высоком соотношении
нерестовых и вырастных площадей в р. Варзуге численность
271
Таблица 106. Доля карликовых самцов среди самцов-пестряток в популяциях
атлантического лосося на Севере России (наши данные), %
Table 106. Share of precocious males among parr males in the populations of Atlantic salmon
in the North of Russia (our data), %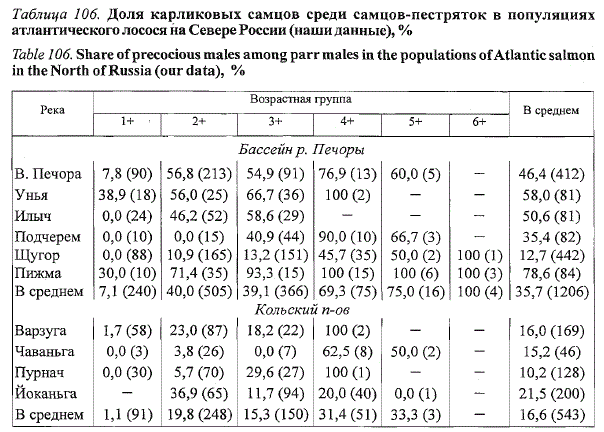
Примечание. В скобках указано количество экземпляров, прочерк - отсутствие данных.
популяции лосося не ограничивается размерами нерестовых участков с
оптимальными условиями для закладки и развития икры, но может лимитироваться
размерами нагульных площадей и уровнем развития кормовой
базы. В результате тугорослость варзугского лосося сформировалась
как приспособление к более эффективному использованию лимитирующих
нагульньж площадей и кормовой базы большим числом особей и
закрепилась на генетическом уровне как полезная адаптация, способствующая
поддержанию численности популяции на высоком уровне.
Небольшие размеры варзугских покатников, равно как и более мелкие
размеры покатников беломорских рек по сравнению с баренцевоморски-
ми в целом, могут быть обусловлены, помимо прочего, возможностью их
промежуточного нагула в опресненных беломорских водах.
Соотношение полов. Сеголетки и годовики лосося характеризуются
равным соотношением полов (см. гл. 4). Преобладание самцов среди стар-
шевозрастных пестряток связано с созреванием части ювенильных самцов
по карликовому типу с последующей задержкой в реке.
На Европейском Севере России в реках, отличающихся более теплым
термическим режимом, часть самцов начинает созревать по карликовому
типу на втором году жизни (табл. 106). Такую картину мы наблюдаем на
НВУ лосося, приуроченных к южной части рассматриваемого региона.
272
В бассейне Печоры в этом возрасте начинают созревать самцы верховий
Печоры (Владимирская, 1957) и р. Уньи (Мартынов, 1983). В притоках
Печоры, расположенных севернее, созревающие самцы начинают встречаться
на третьем (2+) году жизни. На Кольском полуострове карликовые
самцы, созревающие на втором году жизни, отмечены нами в р. Варзуге
(Драганов и др. 1990), которая отличается от других рек полуострова более
высоким содержанием тепла. С возрастом доля созревших самцов
увеличивается, и старшие возрастные группы пестряток могут полностью
состоять из карликовых самцов, например в бассейне Печоры, где
нами встречены восьмилетние (7+) особи. Наличие карликовых самцов,
возраст которых превосходит максимальный возраст покатников в популяции,
является свидетельством их многократного участия в нересте.
Высокая доля карликовых самцов среди пестряток печорского лосося
рассматривается как приспособление к компенсации недостатка крупных
самцов на НВУ (Никольский и др., 1947; Владимирская, 1957). Действительно,
на Европейском Севере России, пожалуй, только в притоках Печоры
старшевозрастные пестрятки (4+ и выше) представлены исключительно
карликовыми самцами. В этом отношении показательна половая структура
пестряток р. Печорской Пижмы, в которой карликовые самцы начинают
созревать на втором году жизни и составляют 100 % в старших возрастных
группах, причем они могут многократно принимать участие в нересте.
Замедление роста карликовых самцов печорского лосося в год созревания
приближает их по этому показателю к туводным видам рыб, рост которых,
как известно, замедляется при созревании.
В популяциях лосося Кольского полуострова (реки Варзуга, Поной,
Йоканьга) самцы на стадии пестрятки принимают участие в воспроизводстве,
как правило, один раз и не образуют контингента многократно
нерестящихся карликовых самцов. Относительно низкая доля карликовых
самцов среди пестряток, по-видимому, связана с отсутствием дефицита
крупных самцов в нерестовых стадах лосося этого региона. В отличие
от печорских у карликовых самцов названных рек рост в год созревания
ускоряется по сравнению с ювенильными ровесниками.
Отнерестившиеся предшествующей осенью карлики встречаются среди
покатной молоди. Их доля среди самцов-покатников в разных популяциях
лосося на Севере России изменяется от 7,0 до 12,0 %. Однако задержка
карликовых самцов в реке, сопровождающаяся повышенной смертностью,
приводит к тому, что во всех популяциях лосося среди мигрирующей
в море молоди преобладают самки, доля которых на несколько процентов
выше, чем самцов.
Созревание пестряток-самцов по карликовому типу определяется наследственными
факторами (Glebe, Saunders, 1986) и модифицируется комплексом
условий среды обитания (Hutchings, Myers, 1994). По данным
Торпа (Thorpe, 1986,1994), созревание пестряток атлантического лосося
по карликовому типу в реках Западной Европы связано с ускорением их
273
роста, т. е. наблюдается та же закономерность, что и в реках Мурмана.
В то же время установлено (Рубан, 1979; Rowe at al., 1991), что при созревании
у карликовых самцов происходит накопление жировых запасов.
Данные А.К. Рубана относятся к печорскому лососю, у которого созревание
самцов по карликовому типу в отличие от рек Мурмана и Западной
Европы сопровождается не ускорением, а замедлением роста. Поэтому
высказанное авторами предположение о том, что толчком к созреванию
ювенильных самцов по карликовому типу является накопление энергетических
запасов до генетически детерминированного уровня представляется
более вероятным, чем ускорение роста.
Карликовые самцы способны не менее эффективно, чем крупные, оплодотворять
икру и оставлять жизнеспособное потомство (Myers, Hutchings,
1987). В период размножения несколько карликовых самцов могут образовывать
совместно с парой крупных рыб так называемые нерестовые
ассоциации. Доля икры, оплодотворенной карликовыми самцами атлантического
лосося в одной из рек Шотландии, в среднем составила 11 %
(Jordan, Youngson, 1992, цит. по: Флеминг, 1998). Карликовые самцы в структуре
проходной формы вида S. salar способствуют повышению устойчивости
популяций, снижая инбридинг, гарантируя оплодотворение икры,
особенно в популяциях, где среди нерестовых мигрантов преобладают
самки.
Катадромная миграция молоди в реках. Длины лососевых рек первого
порядка на Севере России колеблются от 39 км (р. Сидоровка) до
1800 км (реки Печора, Северная Двина с Вычегдой) и охватывают практически
весь спектр изменчивости данного показателя в пределах ареала.
Большие различия в протяженности лососевых рек послужили основанием
для предположения о соответствующих различиях в продолжительности
ската молоди лосося в них (Владимирская, 1957; Дирин, 1981,
19856; см. гл. 4). Скат молоди из рек в море приурочен к периоду с благоприятными
для мигрантов условиями, который на Севере России крат-
ковременен (Бакштанский и др., 1981; Мартынов, 1983; Кузьмин, 1985;
Неклюдов, 1985; Лоенко, Черницкий, 1986; Веселов, Каможин, 2001; и
др.). Средняя продолжительность покатнои миграции смолтов лосося на
Севере России независимо от длины реки равняется одному месяцу с отклонениями
в несколько дней в ту или иную сторону. Скат основного
контингента молоди проходит в еще более сжатые сроки и укладывается
в две недели.
Относительно короткий период ската молоди лосося в притоках
р. Печоры (Мартынов, 1983) может рассматриваться как адаптация к миграции
в протяженных реках, где «миграционное окно» по мере прогрева
воды перемещается в магистральном русле от верховьев к устью, а в притоках
- от устья к верховьям (см. гл. 4). В популяциях, воспроизводящихся
в таких крупных реках, как Печора, видимо, имеет место осенняя предпо-
катная миграция пестряток лосося из горной части нерестовых притоков
в равнинную (в пределах нерестового района). Данное явление послужило
дополнительным основанием для предположения о растянутости
покатной миграции молоди лосося в крупных речных системах и зимовки
смолтов в магистральном русле основной реки (Дирин, 1981). На наш
взгляд, предпокатная миграция старшевозрастных пестряток из верховий
в нижнее течение нерестовых притоков Печоры может рассматриваться
как популяционное приспособление, позволяющее большей части рыб
попасть в «миграционное окно».
Характер миграции покатников определяется комплексом условий
(Бакштанский и др., 1976; Лоенко, Черницкий, 1986; Варнавский, 1990),
среди которых абиотические рассматриваются как фактор, запускающий
и координирующий процессы смолтификации и миграции рыб (Баранникова,
1975). В период покатной миграции в качестве основного фактора,
во многом модифицирующего остальные, выступает температура воды,
которая определяет не только начало и динамику, но и трофические условия
ската через воздействие на развитие биоты водоемов.
По календарным срокам миграция смолтов лосося в северных реках
России приходится на июнь-июль и во всех исследованных популяциях
укладывается в один месяц. Вариации сроков ската не выходят за пределы
июня-июля и определяются межгодовыми и локальными особенностями
гидрометеорологических условий.
Катадромная миграция пестряток атлантического лосося для нагула в
эстуарной зоне достаточно широко распространена в реках северных провинций
Канады (Power, 1969; Power, Shooner, 1966;Hutchings, 1986; Cunjak,
Chadwick, 1989). Основная часть пестряток мигрировала в эстуарий из
маленькой лососевой реки на северо-западном побережье Ньюфаундленда
весной за неделю до ската смолтов и лишь небольшая часть осенью
(Cunjak, Chadwick, 1989). Покатники покидали эстуарий, а пестрятки нагуливались
там в течение лета и возвращались на зимовку в реку. На севере
России катадромная весенняя миграция пестряток лосося отмечена в
беломорской р. Стрельне (Долотов, 1997). По устному сообщению
Э.Л. Бакштанского, мелкая покатная молодь лосося р. Варзуги по своим
характеристикам ближе к пестрятке, чем к смолту. Ее смолтификация, по-
видимому, завершается в распресненных водах Белого моря, которые используются
покатной молодью лосося и других беломорских рек для промежуточного
нагула. Пребывание постсмолтов лосося в Белом море отражается
на чешуе части взрослых рыб в виде переходного кольца.
На северо-востоке ареала постсмолты атлантического лосося из рек,
впадающих в Баренцево море, перемещаются к районам нагула, расположенным
в Норвежском море, против общего направления теплых квазистационарных
морских течений. Молодь лосося из беломорских рек до
выхода в открытый океан определенное время проводит в водах Белого
моря и, по-видимому, попадает в Баренцево море, придерживаясь стоковых
течений. Постсмолты лосося появляются в западной части Баренце-
ва моря на границе с Норвежским в октябре-ноябре, о чем свидетельствует
повышение вылова в это время на единицу усилия ярусами за счет
рыб возраста Р+0+ (Шестопал, Мартынов, 1985).
Смотрите также