РОСТ МОЛОДИ АТЛАНТИЧЕСКОГО ЛОСОСЯ НА СЕВЕРО-ВОСТОКЕ АРЕАЛА
Соотношение линейного и весового роста молоди. В связи с выраженной
сезонной изменчивостью количества резервных веществ, особенно
у молоди атлантического лосося в речной период жизни, характеристики
линейно-весового соотношения, полученные до и после зимовки
рыб, должны различаться, что мы и наблюдаем на примере пестряток
щугорского лосося (рис. 76). Рост молоди в этом уральском притоке Печоры
начинается в первой половине июня и завершается в конце августа.
За время зимовки, продолжительность которой достигает 9 мес, масса
одноразмерных рыб снижается во всех возрастных группах. Подставив в
уравнения регрессии, отражающие линейно-весовое соотношение в конце
и начале нагульного сезона, средние значения длин рыб в возрастных
классах, получим, что за время зимовки масса годовиков (1 •) снижается
на 13 %, двухгодовиков (2-) - на 19 %, трехгодовиков (3-) - на20 %, четы-
рехгодовиков (4-) - на 24 %. С возрастом наблюдается увеличение потерь
массы тела пестряток за зимний период. Регрессия массы тела происходит
за счет уменьшения запасов питательных резервных веществ, о чем
свидетельствует снижение жирности пестряток весной по сравнению с
прошедшей осенью (Рубан, 1979).
При обратных расчислениях роста рыб мы оперируем показателями
их длин в конце нагульного сезона. Расчисленным показателям длин должны
соответствовать показатели массы рыб осенью перед зимовкой.
Поэтому оценивать линейно-весовое соотношение пестряток следует по
материалам, собранным в конце нагульного сезона. Пригодны также материалы,
собранные в августе, желательно в его второй половине. Близкие
значения коэффициентов уравнений регрессии линейно-весового
соотношения у молоди в исследованных нами популяциях (рис. 77) свидетельствуют
о незначительной межпопуляционной изменчивости дан-
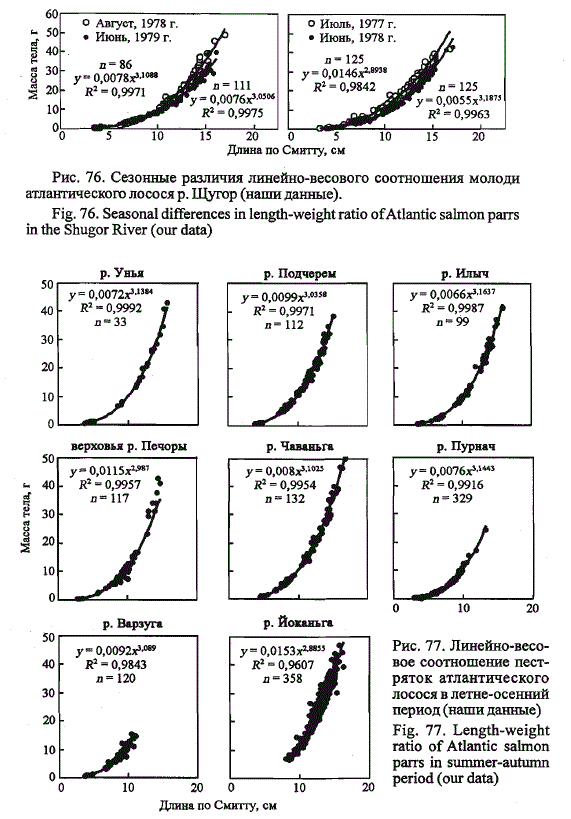
Рис. 76. Сезонные различия линейно-весового соотношения молоди
атлантического лосося р. Щугор (наши данные).
Fig. 76. Seasonal differences in length-weight ratio of Atlantic salmon parrs
in the Shugor River (our data)
Рис. 77. Линейно-весовое
соотношение пест-
ряток атлантического
лосося в летне-осенний
период (наши данные)
Fig. 77. Length-weight
ratio of Atlantic salmon
parrs in summer-autumn
period (our data)
ного показателя у атлантического лосося на северо-востоке ареала в речной
период жизни. Однако при ретроспективной оценке весового роста
целесообразно пользоваться популяционными уравнениями регрессии.
Межгодовые особенности роста пестряток. Прежде чем сравнивать
рост молоди лосося в реках, следует рассмотреть его внутрипопуляцион-
ные особенности. В большинстве случаев сбор материала в полевых условиях
приходится на период активного роста рыб. Полученные при этом
данные отражают не конечные годовые, а промежуточные показатели размерно-
весовых характеристик, что затрудняет количественную оценку различий
роста рыб из разных популяций. Обратное расчисление роста рыб,
даже при наличии неизбежных в этом случае погрешностей, позволяет
получить более близкие к значениям годовых приростов характеристики.
Рост атлантического лосося в речной период жизни подвержен значительной
внутрипопуляционной изменчивости, которая проявляется в
большом размахе колебаний размерно-весовых показателей одновозрас-
тных рыб (Никольский и др., 1947; Владимирская, 1957; Смирнов, 1971;
Мартынов, 1983; Шустов, 1983; Митанс, 1975; и др.). Вариация длины и
массы пестряток с возрастом снижается. Более высокая изменчивость
размеров младшевозрастных пестряток связана с присутствием среди них
как быстрорастущих и впоследствии рано скатывающихся особей, так и
тугорослых рыб с поздним скатом в море.
Рост молоди в уральских притоках Печоры начинается и завершается
при прогреве и охлаждении воды до 7 °С (Мартынов, 1983). Для рек Карелии
и Прибалтики пороговые температуры роста молоди лосося указываются
в пределах 10 °С (Смирнов, 1971; Митанс, 1975). Более низкие
значения температурных порогов роста в холодноводных реках Урала следует
рассматривать как приспособление к существованию вида в более
суровых условиях обитания. Кроме того, временная разница между прогревом
и охлаждением воды в интервале от 7 до 10 °С невелика и, как
правило, не превышает одной недели, а в годы с интенсивным весенним
прогревом и осенним охлаждением воздуха составляет несколько дней.
С учетом пороговых температур продолжительность периода роста молоди
атлантического лосося в северных реках России достигает 3 мес. По
нашим данным, продолжительность роста молоди имеет возрастные особенности,
которые состоят в том, что раньше начинается и позже завершается
рост младшевозрастных пестряток. В весенний период временные
различия между началом роста пестряток разного возраста незначительны.
Осенью разница между завершением роста сеголеток и старше-
возрастных пестряток может достигать двух, а то и трех недель. Особенно
отчетливо указанная выше тенденция проявляется в годы с теплым
летом. Так, в 1977 г. узкие склериты по периметру чешуи у некоторых
старшевозрастных пестряток р. Щугор начинали закладываться в середине
августа, в то время как сеголетки имели «открытый край» чешуи до
конца первой декады сентября.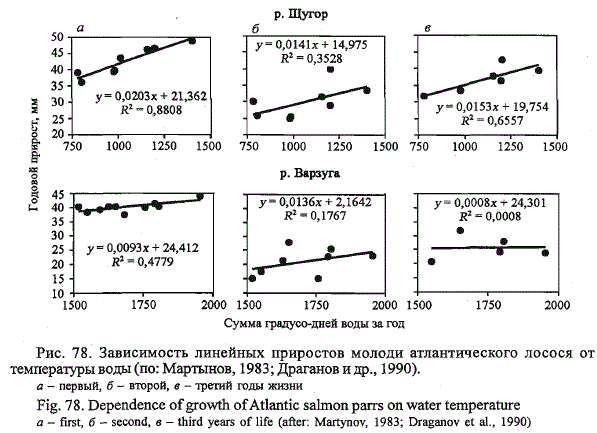
Рис. 78. Зависимость линейных приростов молоди атлантического лосося от
температуры воды (по: Мартынов, 1983; Драганов и др., 1990).
а - первый, б - второй, в - третий годы жизни
Fig. 78. Dependence of growth of Atlantic salmon parrs on water temperature
a - first, б - second, в - third years of life (after: Martynov, 1983; Draganov et al., 1990)
Между ростом молоди лосося и температурой воды в нерестовых реках
существует положительная зависимость (Мартынов, 1983; Драганов
и др., 1990). Наиболее тесная связь между приростами длины и суммой
тепла воды наблюдается в первый год жизни молоди (рис. 78). С возрастом
данная связь ослабевает, о чем свидетельствует снижение значений
коэффициентов корреляции. Более высокая межгодовая зависимость роста
сеголеток от суммы тепла воды, по-видимому, обусловлена особенностями
занимаемых стаций. В лососевых реках сеголетки приурочены к
прибрежью, где за счет инсоляции в теплые солнечные годы вода прогревается
более интенсивно по сравнению с русловой частью, что приводит
к увеличению размаха межгодовых колебаний суммы тепла именно
на стации, занимаемой сеголетками. Кроме того, в холодные дождливые
годы стация сеголеток более подвержена негативным последствиям
повышенной изменчивости уровенного режима.
В холодноводньгх реках (Щугор) рост молоди по сравнению с тепло-
водными (Варзуга) в большей мере зависит от межгодовых колебаний
теплосодержания вод. По-видимому, с этим связана низкая межгодовая
изменчивость средних показателей длины и массы покатников варзуг-
ского лосося (см. гл. 3).
Половые особенности роста пестряток. На нерестилищах пестрят-
ки представлены ювенильными самками, ювенильными и карликовыми
Таблица 88. Годовые приросты длины тела (над чертой - мм) и число склеритов (под
чертой) на чешуе пестряток атлантического лосося р. Подчерем (наши данные)
Table 88. Annual increases of body length (above line) and number of scleritis (under line) on
scale of Atlantic salmon parr in the Podcherem River (our data)
Примечание. Рыба отловлена неводом ячеей 4 мм в приводе с 19.08 по 24.08. 1979 г.
самцами. Анализ результатов прямых наблюдений за ювенильными пест-
рятками (см. выше) свидетельствует об отсутствии половых различий в их
размерно-весовых показателях и подтверждает имеющиеся в литературе.
Во всех популяциях часть пестряток-самцов созревает по карликовому
типу. Выше на примере лосося рек Печоры, Варзуги, Йоканьги и По-
ноя показано, что карликовые самцы по размерам отличаются от своих
ювенильных ровесников, причем в притоках Печоры их длина и масса
достоверно ниже, а в реках Кольского п-ова выше, чем у неполовозрелых
ровесников. Анализ роста двух групп самцов свидетельствует об отсутствии
различий в их приростах вплоть до начала созревания. В бассейне
Печоры созревание карликов сопряжено с замедлением их роста. Так, в
р. Подчерем абсолютные значения приростов в год созревания у карликовых
самцов оказались в среднем на 26,9 % ниже, чем у ювенильных, а
на их чешуе откладывалось на 1,8 склерита меньше (табл. 88). Аналогичная
картина наблюдается в других уральских притоках Печоры. В реках
Кольского полуострова созревание самцов по карликовому типу сопровождается
ускорением роста (см. табл. 67, 69, 81).
Таким образом, на Севере России созревание самцов лосося по карликовому
типу в одних популяциях сопровождается замедлением, а в других
- ускорением роста. В первом случае характер роста подобен большинству
туводных видов рыб (Мина, Клевезаль, 1976), а во втором может
быть классифицирован как пубертатный. По нашим данным, ускорение
роста карликовых самцов при созревании характерно для популяций,
воспроизводящихся в небольших реках, относительно недалеко отстоящих
от морских районов нагула (Кольский п-ов), а замедление - в популяциях
протяженной речной системы (р. Печора), более чем на 2 тыс. км
удаленной от основных районов нагула в море. В первой группе популя-
Таблица 89. Доля рыб с «закрытым» краем речной зоны чешуи среди производителей и
покатников атлантического лосося в реках Севера России (наши данные), %
Table 89. The share offish with a «closed» edge in a river zone of scales in adult and smolt
Atlantic salmon from the rivers of North Russia (our data), %
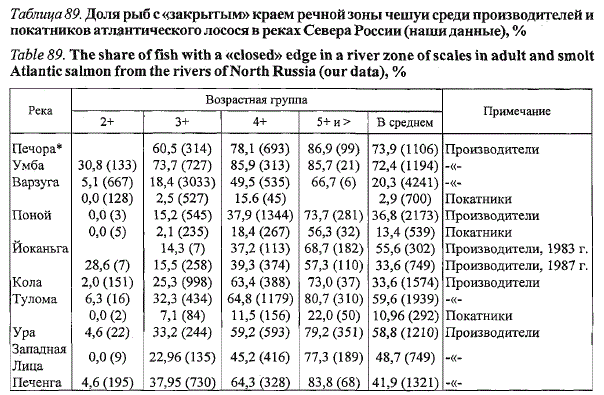
Примечание. В скобках указано количество экземпляров; * — рыба выловлена в устье главной реки.
ций возрастной ряд карликовых самцов укладывается, а во второй выходит
за пределы колебаний речного возраста анадромных мигрантов. Данное
обстоятельство свидетельствует о преимущественно однократном созревании
ювенильных самцов в реке в первом случае и об их повторном
созревании - во втором.
Рост покатников в период катадромной миграции. Скат молоди атлантического
лосося в море непродолжителен по времени и укладьюается
в один месяц, а массовая миграция продолжается не более двух недель.
В этот период часть молоди растет (Неклюдов, 1987,1990; Долотов, 1997).
Речные приросты в год ската в море имеются на чешуе у той или иной
части рыб во всех исследованных нами популяциях.
В некоторых реках индивидуальные приросты покатников в год ската
могут достигать значений годовых приростов на стадии пестрятки. Соотношение
рыб с «закрытым» и «открытым» краем чешуи у покатников в
разных северных реках России изменяется от доминирования первых в
Печоре и Умбе до преобладания последних в реках Мурмана (табл. 89).
Соответствующим образом изменяются и абсолютные значения приростов
длины тела, причем положительная связь между абсолютными значениями
приростов в год ската и длиной рек отсутствует. Наоборот, в
самой протяженной речной системе (р. Печора) наблюдается наименьшая
доля покатников с приростами на чешуе и их абсолютными значениями,
а в коротких реках Мурмана данные показатели достигают макси-
мальных значений. Во всех популяциях среди младшевозрастных покат-
ников больше доля особей с «открытым» краем чешуи и большие значения
абсолютных приростов. Максимальные приросты имеют покатники
самых младших возрастных групп, а минимальные - рыбы старшей возрастной
группы (Неклюдов, 1990; Черницкий, Лоенко, 1990; Долотов,
1997; наши данные). Таким образом, возрастные особенности роста в
период ската приводят к сглаживанию различий между средними размерами
разновозрастных покатников. Средние показатели длины и, следовательно,
массы у покатников крайних возрастных групп существенно
сближаются. Выводы, полученные на основе анализа материалов обратного
расчисления роста по чешуе производителей, подтверждаются прямыми
наблюдениями за мигрирующей в море молодью.
Популяционные особенности роста молоди лосося в период покат-
ной миграции состоят в различной степени выраженности рассмотренного
выше феномена, крайние варианты которого демонстрируют материалы
по рекам Печора и Йоканьга (см. гл. 7).
Межпопуляционные особенности роста. Сопоставление годовых
приростов одновозрастных покатников из исследованных нами популяций
лосося выявило существенные межпопуляционные различия между
ними (рис. 79, табл. 29-41 Приложения). Можно выделить две группы
популяций, различающиеся по характеру роста в речной период жизни.
Более высокие показатели роста у лосося рек, впадающих в Баренцево
море. Среди них во всех возрастных группах по темпу роста в речной
период жизни доминирует лосось р. Йоканьги. Для беломорских рек характерен
более низкий темп речного роста. Наименьшими показателями
роста характеризуется варзугский лосось.
Рост молоди лосося в экспериментальных условиях. В естественных
условиях рост молоди атлантического лосося имеет выраженный
ритмический характер: интенсивный прирост длины и массы тела летом
сменяется приостановкой роста и снижением массы тела зимой. Периодичность
роста молоди определяется сезонными колебаниями климатических
условий, а его скорость — суммой тепла, аккумулированного тем
или иным водным объектом в конкретных физико-географических условиях,
и наследственными особенностями популяций. На рыбоводных
заводах для интенсификации роста молоди лосося в качестве одного из
методов используется подогрев воды (Яндовская, Казаков, 1974; Яндов-
ская и др., 1979), позволяющий получать покатную молодь в течение одного
года вместо двух-трех в природе. При этом температурный режим
существенно отличается от такового в естественных условиях проживания
выращиваемого объекта. Изучение особенностей роста «дикой» молоди
лосося в эксперименте представляет интерес для оценки эндогенных
и экзогенных факторов роста. Такая возможность представилась при
организации экспериментального выращивания молоди лосося р. Колы
на Кислогубском экспериментальном участке ПИНРО (1984 - 1985 гг.) и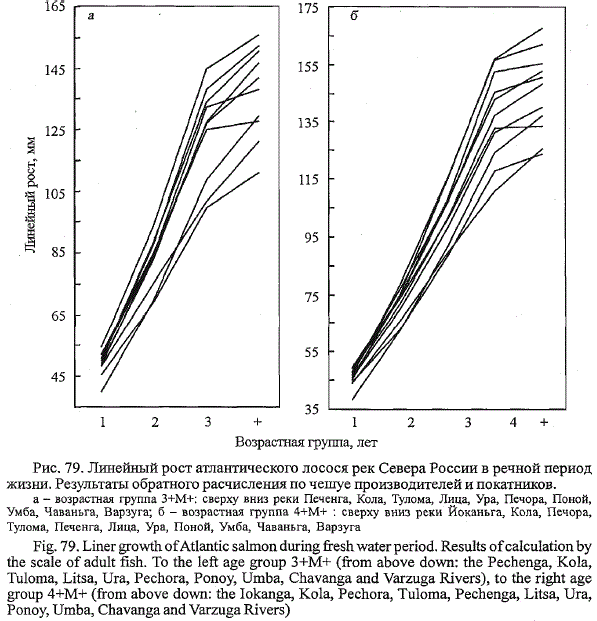
Рис. 79. Линейный рост атлантического лосося рек Севера России в речной период
жизни. Результаты обратного расчисления по чешуе производителей и покатников.
а - возрастная группа 3+М+: сверху вниз реки Печенга, Кола, Тулома, Лица, Ура, Печора, Поной,
Умба, Чаваньга, Варзуга; б - возрастная группа 4+М+ : сверху вниз реки Йоканьга, Кола, Печора,
Тулома, Печенга, Лица, Ура, Поной, Умба, Чаваньга, Варзуга
Fig. 79. Liner growth of Atlantic salmon during fresh water period. Results of calculation by
the scale of adult fish. To the left age group 3+M+ (from above down: the Pechenga, Kola,
Tuloma, Litsa, Ura, Pechora, Ponoy, Umba, Chavanga and Varzuga Rivers), to the right age
group 4+M+ (from above down: the Iokanga, Kola, Pechora, Tuloma, Pechenga, Litsa, Ura,
Ponoy, Umba, Chavanga and Varzuga Rivers)
Верхнетуломском рыбоводном заводе (Мартынов, Горшкова, 1986; Мартынов
и др., 1998).
Температура воды в период экспериментальных работ на Кислогубс-
ком участке колебалась от 5 до 18 °С, на Верхнетуломском рыбоводном
заводе - от 6 до 15 °С (рис. 80). На протяжении почти всего периода
вьфащивания молоди она превышала 10 °С и приближалась к оптимальным
значениям для вида. Понижения температуры воды имели место во
второй половине декабря и первой половине января на Кислогубском
экспериментальном участке и были непродолжительны по времени.
На Кислогубском экспериментальном участке молодь выращивали 328
дней, на Верхнетуломском заводе - 313. Расхождение календарных сро-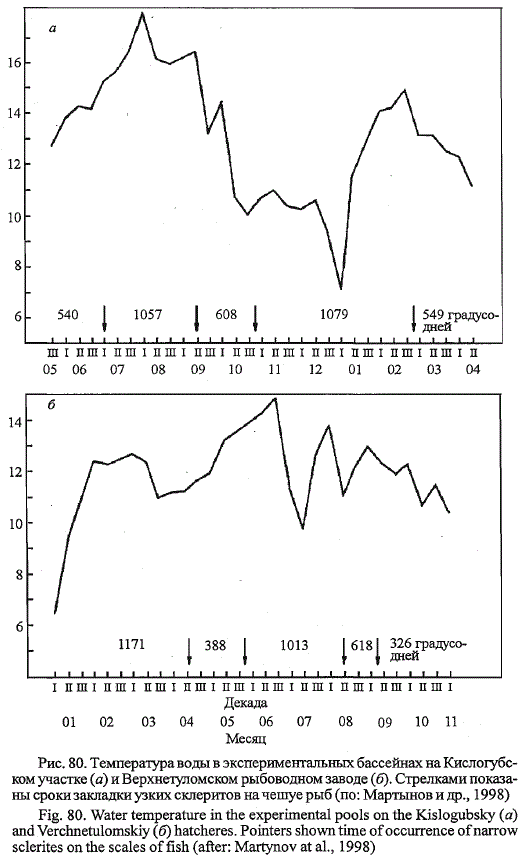
Рис. 80. Температура воды в экспериментальных бассейнах на Кислогубс-
ком участке (а) и Верхнетуломском рыбоводном заводе (б). Стрелками показаны
сроки закладки узких склеритов на чешуе рыб (по: Мартынов и др., 1998)
Fig. 80. Water temperature in the experimental pools on the Kislogubsky (a)
and Verchnetulomskiy (6) hatcheres. Pointers shown time of occurrence of narrow
sclerites on the scales offish (after: Martynov at al., 1998)
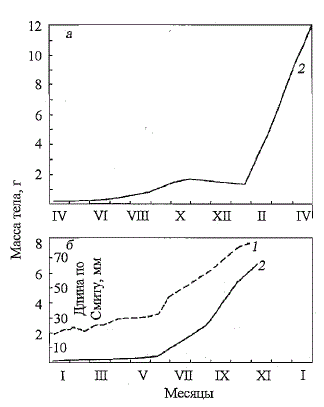
Рис. 81. Линейный (1) ивесовой (2)
рост молоди атлантического лосося на
Кислогубском участке (а) и Верхнету-
ломском рыбоводном заводе (б) (по:
Мартынов и др., 1998)
Fig. 81. Liner (1) and weight (2)
growth of Atlantic salmon parrs on the
Kislogub-sky (a) and Verchnetulomskiy
(6) (after: Martynov at al., 1998)
ков выращивания рыб объясняется
выклевом личинок в разные
месяцы, так как в первом
случае икра инкубировалась
при естественном температурном
режиме, а во втором применялся
подогрев воды.
К концу периода выращивания
средняя масса молоди на
Кислогубском участке составила
12,1 г при средней длине по
Смитту 120 мм, а на Верхнету-
ломском рыбоводном заводе соответственно 6,53 г и 79 мм. Наибольшие
приросты массы и длины рыб наблюдались во второй половине экспериментальных
работ (рис. 81). На Кислогубском экспериментальном участке
рост рыб приостанавливался зимой с ноября 1984 г. по конец января
1985 г., несмотря на положительные температуры воды (от 5 до 14 °С),
полноценные рационы и хорошую поедаемость рыбами корма. В этот
период масса тела рыб снижалась, в то время как средние показатели длины
находились на одном уровне.
Во избежание негативных последствий от частых манипуляций с рыбами
контрольное взвешивание и измерение осуществляли через относительно
длительные промежутки времени, вследствие чего кривые роста
имеют сглаженный характер и не отражают особенности роста за короткие
промежутки времени. В связи с этим для изучения ритмики роста рыб
нами были проанализированы сроки и условия закладки на чешуе рыб
узких склеритов, отражающих замедление их роста (Мартынов, Горшкова,
1986; Мартынов и др., 1998).
На Кислогубском участке на чешуе молоди выявлено пять зон роста,
разграниченных четырьмя кольцами из одного-трех узких склеритов. Первое
кольцо формировалось у 49 % рыб при средней длине тела 32 см
(табл. 90) в первой декаде июля, когда суточные значения температуры
воды колебались в пределах 13-15 °С. Второе кольцо было четко выражено
на чешуе всех исследованных рыб и закладывалось в середине сентября
при температуре воды 15 °С по достижении рыбами средней дли-
Таблица 90. Число склеритов в зонах роста чешуи и расчисленные длины молоди
атлантического лосося р. Колы, выращенной на Кислогубском экспериментальном
участке (по: Мартынов и др., 1998)
Table 90. The number of sclerites in the scale zones and back calculated length of young
Atlantic salmon from the Kola River reared on the experimental farm of Kislogubsky hatchery
(after: Martynov et al., 1998)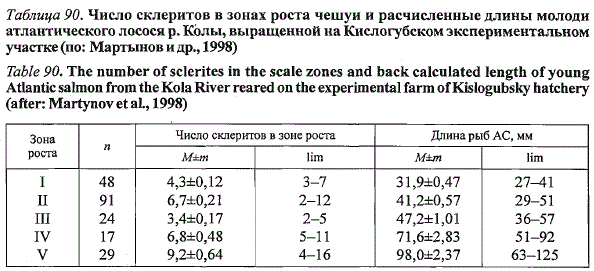
ны 42 мм. После кратковременной приостановки в сентябре рост рыб
возобновился и продолжался до конца октября. В конце октября - начале
ноября рост рыб прекратился в результате снижения температуры воды и
возобновился лишь в начале января. Остановка роста молоди лосося в
ноябре-декабре сопровождалась закладкой узких склеритов, сформировавших
у 73 % рыб третье дополнительное кольцо. И, наконец, четвертое
кольцо из одного-двух узких склеритов образовалось на чешуе экспериментальных
рыб в конце февраля-начале марта при температуре воды
12—13 °С при средней длине 69 мм.
Аналогичная ритмика роста молоди Кольского лосося наблюдалась в
условиях повторного эксперимента на Верхнетуломском рыбоводном заводе
(табл. 91). Длины рыб в конце формирования зон суженых склеритов
оказались близкими в условиях обоих экспериментов, несмотря на
различия в календарных сроках проведения работ, температурном режи-
Таблица 91. Число склеритов в зонах роста чешуи и расчисленные длины молоди
атлантического лосося р. Колы, выращенной на Верхнетуломском рыбоводном заводе
(по: Мартынов и др., 1998)
Table 91. The number of sclerites in the scale zones and back calculated length of young
Atlantic salmon of the Kola River reared on the Upper Tuloma hatchery (after: Martynov at al.,
1998)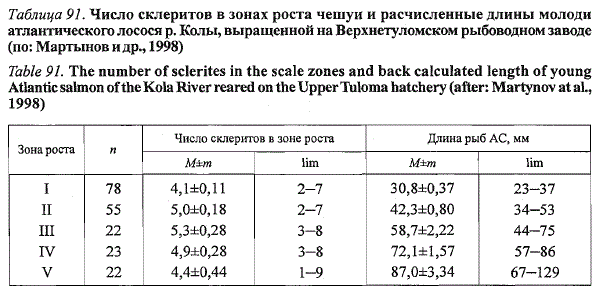
Таблица 92. Число склеритов в годовых зонах роста чешуи и расчисленные длины «диких»
пестряток атлантического лосося р. Колы (по: Мартынов и др., 1998)
Table 92. The number of sclerites in the annual zones of scale and back calculated length of
«wild» Atlantic salmon parr in the Kola River (after: Martynov et al., 1998)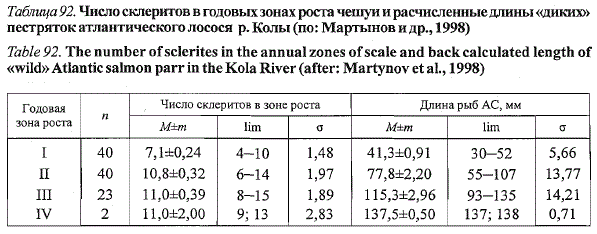
ме инкубации икры и выращивания молоди. В обоих случаях на чешуе
молоди можно было выделить пять зон роста, разграниченных четырьмя
кольцами из суженных склеритов. В большинстве случаев зоны узких склеритов,
показывающие замедление роста рыб, формировались при высоких
температурах воды.
Не исключено, что первое кольцо отражает замедление роста, связанное
с морфогенезом при переходе от личиночной стадии к мальковой.
Аналогичные кольца могут формироваться на чешуе части рыб в естественных
условиях (см. гл. 2). Второе кольцо формируется у всех экспериментальных
рыб при длине, соответствующей длине годовиков Кольского лосося
в естественных условиях (табл. 92), третье — лишь у части особей и,
видимо, является дополнительным, отражая замедление роста рыб при
понижении температуры воды в эксперименте. Интересно, что четвертое
кольцо, как и второе, формировалось у всех экспериментальных рыб при
длине, соответствующей длине двухгодовиков в естественных условиях.
Годовые приросты молоди лосося, особенно в первый год жизни,
положительно коррелируют с суммой градусо-дней воды (Мартынов,
1983; Драганов и др., 1990). На Кислогубском экспериментальном участке
ко времени образования второго кольца она составила 1597 градусо-
дней, а за период роста рыб от времени закладки второго до формирования
четвертого кольца - 1673. При повторном эксперименте в 1986 г. на
базе Верхнетуломского рыбоводного завода за периоды формирования соответствующих
колец эта сумма составила 1559 и 1631 градусо-дней.
По данным службы ГМС (1429-й км Октябрьской железной дороги),
среднегодовая температура воды в р. Коле составила 1529,2±33,2 градусо-
дней (п = 25 лет) при годовых колебаниях от 1170 до 1802. Совпадение
суммы тепла, по достижении которой у экспериментальных рыб происходила
закладка второго и четвертого колец, со средней многолетней
суммой тепла в их «родной» реке позволяет предположить, что в процессе
исторического становления в популяции Кольского лосося сложился
генетически закрепленный тип циклов развития молоди, «притертый» к
212
конкретным абиотическим условиям обитания. В этом случае замедление
роста рыб в эксперименте при благоприятных для роста внешних
условиях при достижении ими характерных для дикой популяции линейно-
весовых параметров в конце нагульного сезона можно объяснить наследственной
обусловленностью цикличности онтогенеза.
В отличие от южных районов ареала, например рек Прибалтики, где
средний возраст покатной молоди составляет 1,8 года (Митанс, 1975), в
северных реках длительность речного периода жизни лосося значительно
выше - до 4,7 года и более (Кузьмин, Смирнов, 1982; Мартынов и др.,
1988; Power, 1969; и др.). Соответственно на севере ареала молодь в своем
развитии до покатного состояния проходит большее число годовых
циклов развития, характеризующихся чередованием периодов интенсивного
роста летом и его приостановки в зимние месяцы.
В северных реках продолжительность роста молоди лосося, как правило,
не превышает 3 мес. (с середины июля до середины сентября). Сеголетки
лосося р. Мирамичи (Канада) предпочитают температуру около
17 °С (Javaid, Anderson, 1967). Наиболее интенсивный рост молоди наблюдается
в интервале температур 13-16 °С (Dwyer, Piper, 1987). Если
выращивать Кольскую молодь лосося при таких температурах, то для набора
среднегодовой суммы тепла, характерной для р. Колы (1529 граду-
со-дней), потребуется примерно три месяца, т. е. за этот отрезок времени
в экспериментальных условиях могут быть получены среднегодовые приросты
длины и массы рыб, характерные для популяции в естественных
условиях.
В естественных условиях рост молоди лосося в конце нагульного периода
прекращается при снижении температуры воды до 7 °С (Allen, 1969;
Мартынов, 1983). При выращивании молоди периоду относительного
физиологического покоя рыб, сопровождающегося приостановкой их роста,
должна соответствовать пониженная температура воды. В естественных
условиях она близка к О °С. Хотя обоснование температурного режима
и длительности периода покоя при искусственном выращивании рыб
требует дополнительных исследований, очевидно, что при ускоренном
выращивании холодноводных рыб следует учитывать популяционные
особенности годовой ритмики их роста.
Ускоренное прохождение наследственно детерминированных годовых
циклов роста приводит к сокращению хронологического возраста рыб, в
то время как их биологический возраст не изменяется. У большинства
акселерированной молоди Кольского лосося второе и четвертое кольца на
чешуе, скорее всего, отражают их биологический возраст (2+ по аналогии
с «дикой» молодью), в то время как их хронологический возраст соответствует
одному году (1-)- По-видимому, аналогичное толкование было бы
уместно при объяснении расхождения хронологического (1-) и определенного
по чешуе (4-) возраста покатника атлантического лосося, выращенного
в заводских условиях в Норвегии (Shearer, 1989, plate 31).
Смотрите также