
ЭКОЛОГИЯ АНАДРОМНЫХ МИГРАНТОВ АТЛАНТИЧЕСКОГО ЛОСОСЯ В РЕКАХ СЕВЕРА РОССИИ
Созревание. В реке происходят созревание половых продуктов взрослых
рыб и их размножение. После захода в реку гонады рыб летней биологической
группы созревают в течение 1,5 — 3,5 мес, в то время как у
«озимых» рыб - 11,5 - 13,5 мес. Лосось при заходе из моря в реки имеет
слабо развитые половые железы. Средние показатели коэффициентов
зрелости «ярового» лосося Западного Мурмана при заходе в реки примерно
в три раза выше, чем у «озимых» рыб р. Печоры (табл. 63). Коэффициенты
зрелости «озимых» самок при заходе в р. Онегу колеблются от
0,35 до 0,67 % (Конрадт, 1949). Более высокие коэффициенты зрелости
«ярового» лосося р. Туломы по сравнению с другими реками Мурмана
связаны с задержкой нерестовых мигрантов в нижнем бьефе Нижнету-
ломской ГЭС. Минимальные значения гонадосоматического индекса «яровых
» рыб перекрываются с максимальными показателями «озимых». Коэффициенты
зрелости мигрирующих на нерест рыб повышаются с увеличением
морского возраста рыб. У самок-тинды они составляют в среднем
0,6 %, в то время как у рыб в возрасте Р+3+ превышают 2,3 %. Коэффициенты
зрелости лосося рек Западного Мурмана увеличиваются к концу
нерестового хода в несколько раз (рис. 62), что свидетельствует о начале
созревания гонад «яровых» рыб в море. Гонады «озимых» рыб при за-
Таблица 63. Коэффициенты зрелости атлантического лосося при заходе из моря в реки
Севера России, %
Table 63. Gonad-somatic indexes of Atlantic salmon entering the rivers in North Russia, %
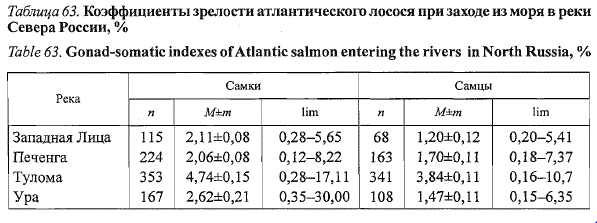
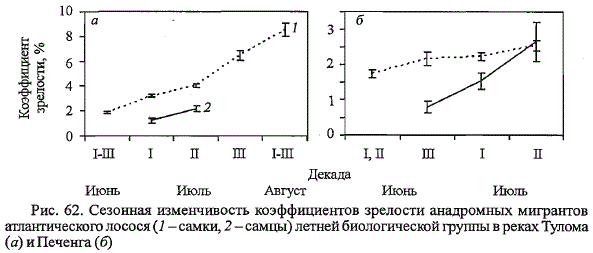
Рис. 62. Сезонная изменчивость коэффициентов зрелости анадромных мигрантов
атлантического лосося (1—самки, 2—самцы) летней биологической группы в реках Тулома
(а)иПеченга(б)
Fig. 62. Seasonal dynamics of gonad-somatic index for «vernal» salmon females (7) and
males (2) entering Tuloma (a) and Pechenga (6) rivers
ходе в реки находятся в стадии покоя, которая длится 9,5 - 11,0 мес. до
весны следующего года (рис. 63).
Нарастание массы гонад «озимых» рыб начинается после вскрытия
рек ото льда, но до конца июня идет очень медленно. Интенсивное формирование
половых продуктов происходит с середины июля до конца
августа. Максимальных значений гонадосоматические индексы рыб достигают
на IV стадии зрелости перед нерестом к середине сентября и в
среднем у «озимых» самок лосося р. Щугор составляют 20,17±3,81 % (и =
= 45 экз.), у самцов - 4,62±1,38 % (п = 8 экз.). Созревание половых продуктов
«яровых» рыб, начавшись еще в море, завершается в пресной воде,
минуя стадию покоя по такой же схеме, что и у «озимых» рыб после зимовки
в реке.
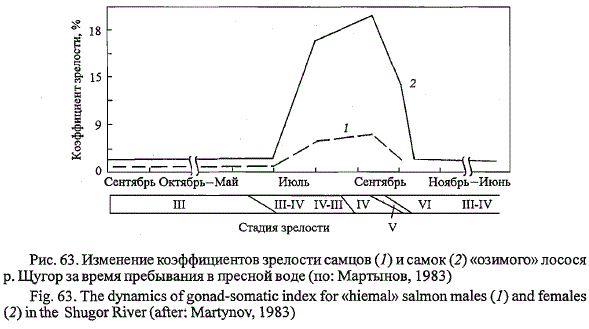
Рис. 63. Изменение коэффициентов зрелости самцов (1) и самок (2) «озимого» лосося
р. Щугор за время пребывания в пресной воде (по: Мартынов, 1983)
Fig. 63. The dynamics of gonad-somatic index for «hiemal» salmon males (/) and females
(2) in the Shugor River (after. Martynov, 1983)
Рис. 64. Снижение массы анадромных мигрантов
атлантического лосося наНВУ р. Щугор по сравнению
с одноразмерными рыбами при заходе в
р. Печору.
Относительные значения массы рыб: 1 - при заходе в
р. Печору; 2 - примерно месяц спустя на нерестилищах р.
Щугор; 3 - после зимовки и созревания половых продуктов;
4- после нереста (по: Мартынов, 1983)
Fig. 64. Loss of weight of adult Atlantic salmon in the Shugor River.
Relative value of weight: 1 - by entering the Pechora River; 2 - one month later in the river Shugor; 3 -
after wintering and maturation; 4 - after spawn (kelts in the Shugor River) (after: Martynov, 1983)
Пребывание взрослых рыб в реке связано с переходом на эндогенное
питание и сопровождается потерями массы тела. Снижение массы рыб
может служить показателем уровня энергетических затрат на том или
ином этапе их пребывания в пресной воде. Рассмотрим динамику изменения
массы тела производителей в крупных речных системах на северо-
востоке ареала на примере «озимого» лосося р. Печоры (рис. 64). Только
при транзитной миграции рыб от устья Печоры до р. Щугор на расстояние
1100 км в течение примерно одного месяца масса рыб снижается на
23,2 %. Затем в течение года, включая зимовку в пресной воде и созревание
половых продуктов, она несколько возрастает, что связано с обводнением
тканей и ростом ооцитов, и вновь резко снижается после
вымета половых продуктов. Потери массы отнерестившихся рыб достигают
23 % от массы зрелых рыб и аналогичны таковым у лосося из других
рек (Warner, 1962; Baum, Meister, 1971). В сравнении с массой одноразмерных
рыб при заходе из моря масса отнерестившихся рыб снижается
на 39,3 %. В процессе посленерестового ската она продолжает падать
и при длительном пребывании вальчаков в реке снижается до 52 %
от первоначальной. Суммарные потери массы печорского лосося составляют
45 - 50 % и сходны с потерями массы у лосося из других рек: р. Во-
лонга- 50 % (Даниленко, 1967), пресноводный лосось из Онежского озера-
40 % (Смирнов, 1971).
Таким образом, 59 % от суммарных потерь массы тела печорского лосося
приходится на транзитную миграцию к нерестилищам и 41 % - на
период формирования половых продуктов и нерест. Несмотря на неизбежные
погрешности при оценке потерь массы рыб путем сопоставления
массы одноразмерных рыб на притоке и в устье главной реки, очевидно,
что более половины потерь приходится на преодоление пресноводного
участка миграционного пути. Данное обстоятельство свидетельствует
о высокой энергетической цене за использование удаленных от
моря репродуктивных участков рек.
На сроки созревания верхнепечорского лосося существенное влияние
оказывает температура воды в летний и осенний периоды: высокая - задерживает,
а низкая - ускоряет созревание гонад (Владимирская, 1960).
Мы такой зависимости для лосося р. Щугор однозначно не выявили. По-
видимому, в условиях такой крупной реки, как Щугор, лосось имеет возможность
избирать места отстоя с благоприятным температурным микрорежимом
даже в годы с теплым летом. По нашим данным, в нерестовых
реках гонады самцов достигают V стадии развития раньше, чем у
самок. Так, на нерестилищах р. Щугор текучие самцы начинают встречаться
примерно на 7 - 10 дней раньше текучих самок. По данным
B.C. Михина (1959а), на нерестилищах северодвинского лосося самцы
появляются раньше самок, что также может быть связано с половыми
различиями в сроках созревания гонад.
Нерест лосося в реках русского Севера начинается в середине сентября
и заканчивается обычно в первой декаде октября (Михин, 1959; Никифоров,
1958,1959а, б; Владимирская, 1957; Мартынов, 1979,1983). В бассейне
Печоры при одинаковой длительности нерестового периода (20 -
22 дня) нерест лосося в североуральском притоке р. Щугор начинается в
среднем на 10 дней и заканчивается на 8 дней раньше, чем в верховьях
Печоры, расположенных южнее (табл. 64), что связано с более ранним
охлаждением воды в р. Щугор. Близкими оказались температурные пороги
начала нереста. Мы, так же как и М.И. Владимирская (1960) для верховий
Печоры, не обнаружили влияния уровенного режима на динамику
нереста щугорского лосося. Его нерест заканчивается, как правило, за несколько
дней до начала ледовых явлений при средней межгодовой температуре
воды 2,2 °С. В верховьях Печоры в отдельные годы лосось завершает
нерест поздней осенью подо льдом (Владимирская, 1960; Кули-
да, 1976а).
Таблица 64. Сроки и температурные пороги (*) нереста атлантического лосося в бассейне
Печоры
Table 64. The terms and temperature thresholds of Atlantic salmon spawning in the Pechora
River basin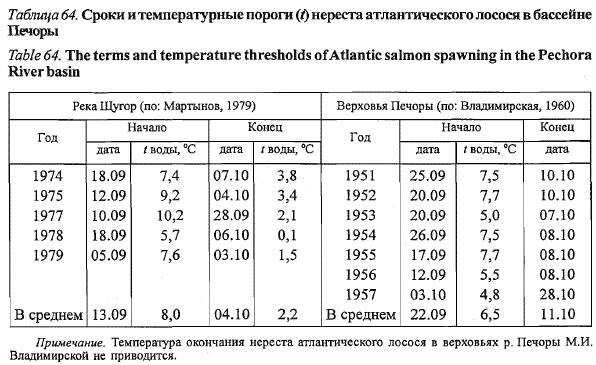
Примечание. Температура окончания нереста атлантического лосося в верховьях р. Печоры М.И.
Владимирской не приводится.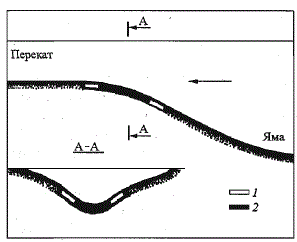
Рис. 65. Типичное расположение
лососевого участка в русле реки.
1 - разреженное расположение бугров; 2 -
наибольшая концентрация бугров (по:
Мартынов, 1979)
Fig. 65. Typical location of salmon reds
in the riverbed.
1 - sparse distribution of reds; 2 - highest
concentration of reds (after: Martynov, 1979)
В руслах таких крупных рек,
как лососевые притоки р. Печоры,
можно выделить два типа
участков, используемых лососем
для закладки нерестовых бугров: стрежневой и прибрежный. В первом,
наиболее типичном случае, икра закладывается на фарватере русла в верхней
части порога в зоне треугольника слива воды (рис. 65). Во втором
для нереста используются отмели прибрежий и островов, расположенные
в зоне порогов. Нерестовые участки «стрежневого» и «прибрежного
» типов различаются механическим составом грунта, скоростями течения
и глубиной воды (табл. 65). Аллювиальный грунт прибрежных отмелей
содержит больше таких мелких фракций, как песок и гравий, что обусловливает
его меньшую проницаемость для подруслового потока. В прибрежье
существенно снижены скорости течений.
На нерестовых участках обоих типов непременным условием является
положительный наклон дна к направлению течения. По-видимому,
именно на таких участках, где надрусловый поток частично переходит в
подрусловый, создаются благоприятные гидрологические условия для
закладки, оплодотворения и развития икры. Как показали наши наблюдения,
откладываемая на этих участках икра не сносится за пределы нерестового
бугра: она «засасывается» потоком в межгалечниковое пространство
нерестового грунта. Кроме того, на входе подруслового потока
развивающаяся на участках стрежневого типа икра гарантирована от
дефицита кислорода в течение всего периода инкубации. Именно на нерестовых
участках стрежневого типа во время осенних ледовых явлений
не формируется донный лед. Закладка нерестовых бугров на участках с
отрицательным наклоном к течению, т. е. на выходе подруслового потока,
нами не отмечена.
По-видимому, именно в зонах частичного перехода надруслового потока
в подрусловый откладывал икру лосось р. Нарвы на неподвижном
хрящеватом грунте, описанном Т.П. Привольневым (1962) при обследовании
нерестилищ, осушенных после строительства Нарвской ГЭС. Приуроченность
нерестовых бугров лосося к русловым участкам, имеющим
положительный наклон к течению, отмечена многими исследователями
(Владимирская, 1957; Михин, Шпайхер, 1957; Михин, 1959а, б; Никифо-
Таблица 65. Характеристика нерестовых участков и размеры нерестовых бугров
атлантического лосося р. Щугор (по: Мартынов, 1979 с дополнениями)
Table 65. Characteristic of spawning grounds and size of redds of Atlantic salmon in the
Shugor River (after: Martynov, 1979 with additions)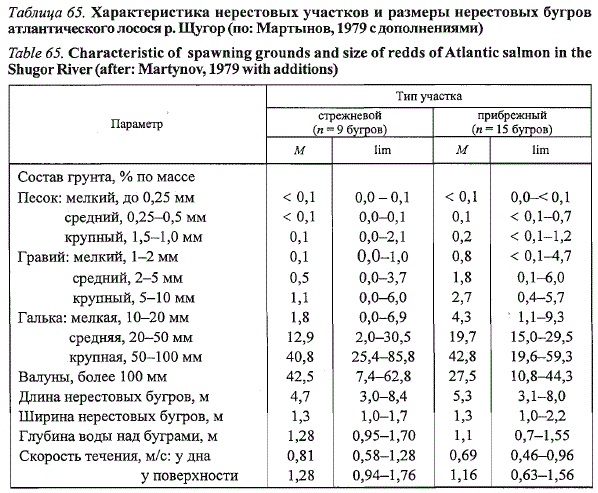
ров, 1958,19596; Мартынов, 1979; Веселов, 1989; Красиков, 1993; Весе-
лов, Калюжин, 2001; и др.). Казалось бы, использование лососем для
откладки икры участков порогов с оптимальными условиями должно гарантировать
высокую эффективность нереста. Однако учет икры в нерестовых
гнездах атлантического лосося демонстрирует противоположные
результаты: до 90 % и более икры может теряться при ее закладке в бугор
(Михин, Шпайхер, 1957; Михин, 1959а; Никифоров, 1959а; Гринюк, 1963).
Данные результаты ставят под сомнение биологическую целесообразность
такого явления в экологии проходных лососей, как забота о потомстве. На
наш взгляд, представление о больших потерях икры лососем во время
нереста, скорее всего, связано с недоучетом икры при ее выемке в условиях
гидродинамического потока. По крайней мере, наша попытка провести
учет отложенной икры на порогах р. Щугор без изоляции нерестового
гнезда от потока воды закончилась неудачей.
Участки с оптимальными гидротопографическими условиями для нереста
и инкубации икры на порогах лососевых рек ограничены по площади.
В р. Щугор нерестовые участки занимают в разные по гидрологическим
условиям годы от 7 до 9 % площади порогов. Флуктуация нерес-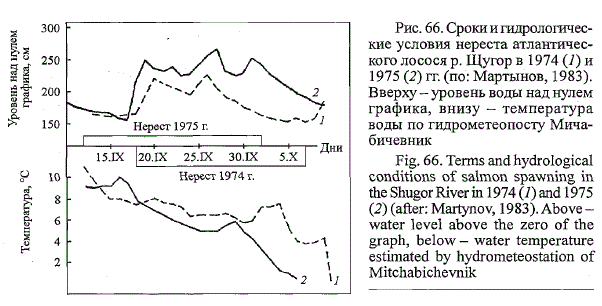
Рис. 66. Сроки и гидрологические
условия нереста атлантического
лосося р. Щугор в 1974 (/) и
1975 (2) гг. (по: Мартынов, 1983).
Вверху - уровень воды над нулем
графика, внизу - температура
воды по гидрометеопосту Мича-
бичевник
Fig. 66. Terms and hydrological
conditions of salmon spawning in
the Shugor River in 1974 (7) and 1975
(2) (after: Martynov, 1983). Above -
water level above the zero of the
graph, below - water temperature
estimated by hydrometeostation of
Mitchabichevnik
товых площадей обусловливается межгодовой изменчивостью уровен-
ного режима воды во время нереста (рис. 66). При подтоплении осенними
дождевыми паводками аллювиальных галечниковых кос на них возникают
благоприятные условия для закладки икры, что провоцирует нерест
рыб на участках прибрежного типа. Так, в 1974 г., когда нерест проходил
при уровнях воды, близких к летней межени, нерестовые бугры
лосося в р. Щугор встречались только на участках стрежневого типа. В
1975 г. при высоких осенних уровнях воды до 40 % бугров были заложены
на затопленных отмелях в прибрежье. Многие из них обсохли при
понижении уровня воды перед ледоставом (Мартынов, 1979).
Таким образом, более благоприятными для выживания икры лосося в
крупных лососевых притоках Печоры являются годы с низким уровнем
воды в период нереста. Ранее к аналогичным выводам при изучении нереста
лосося в верховьях Печоры пришла М.И. Владимирская (1960). Доля
нерестовых участков с оптимальными гидротопографическими условиями
от площади нерестово-вырастных угодий существенно варьирует в
разных лососевых реках. В притоке Печоры р. Щугор она составляет, по
нашим данным, 7 %, в Онеге - 10 % (Кулида, 2002), в Варзуте - 23 %
(Казаков и др., 1992) от площади НВУ.
Большинство нерестовых гнезд лосося на нерестилищах р. Щугор имело
вытянутую овальную форму (рис. 67). По размерам нерестовый бугор,
в который закладывалась икра, многократно превышал нерестовую яму,
формируемую самкой при рыхлении грунта. Глубина нерестовой ямы,
представлявшей собой небольшую покопку, не превышала 10 см, а высота
нерестового бугра в нижней по течению части - 15 см. Высота нерестового
бугра, как правило, возрастала от нерестовой ямы вниз по течению.
В результате угол наклона поверхности нерестового бугра к течению
увеличивался по сравнению с поверхностью грунта. Средняя длина
нерестовых гнезд составляла 4,7 - 5,3 м при средней ширине 1,3 м (см.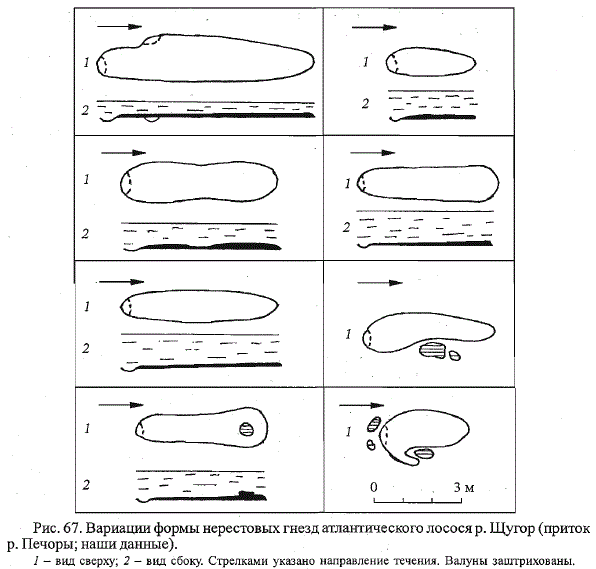
Рис. 67. Вариации формы нерестовых гнезд атлантического лосося р. Щугор (приток
р. Печоры; наши данные).
1 - вид сверху; 2 — вид сбоку. Стрелками указано направление течения. Валуны заштрихованы.
Fig. 67. Form variations of Atlantic salmon redds in the Shugor River (The Pechora River
tributary; our data).
1 - view from above; 2 - view at the side. Current direction is shown by pointers. Boulders are shaded
табл. 65). Отклонения нерестовых гнезд от типичной формы были обусловлены
наличием среди галечникового грунта крупных валунов. В
р. Щугор мы отмечали закладку икры отдельными рыбами в 2 - 3 бугра,
что вызвано их нерестом на участках, где среди галечникового грунта
встречается большое количество валунного материала. Натыкаясь при
строительстве бугра на крупный валун, самка перемещается в сторону и,
продолжая нерест, закладывает новые бугры. Их размеры при этом уменьшались
до 3,0х 1,1 м.
По данным М.И. Владимирской (1957), в верховьях Печоры, где воспроизводится
крупный лосось, большинство нерестовых бугров имели
длину 1,0-1,5 м, а самые большие из них достигали длины 2,0 м, в то
Таблица 66. Механический состав грунта нерестовых бугров атлантического лосося в
верховьях р. Печоры (по: Владимирская, 1957)
Table 66. Mechanical composition of gravel in the Atlantic salmon redds in the upper part of
the Pechora River (after: Vladimirskaya, 1957)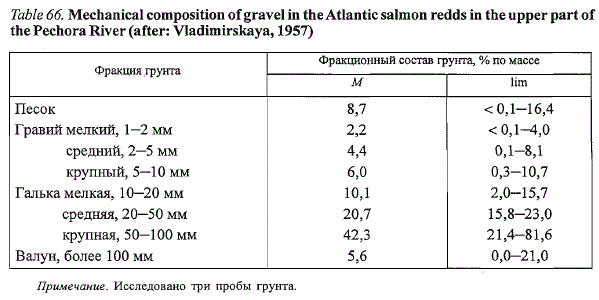
Примечание. Исследовано три пробы грунта.
время как максимальная длина нерестового бугра более мелкого щугор-
ского лосося составила 8,4 м. На наш взгляд, различия в размерах и строении
нерестовых гнезд щугорского и верхнепечорского лосося связаны с
преобладанием в последнем случае мелких фракций грунта на нерестовых
участках (табл. 66). Помимо механического состава грунта, размеры
и строение нерестовых гнезд могут определяться размерами нерестовых
участков. В среднем и нижнем течении р. Щугор, где ширина порожистых
участков достигает 150-200 м, нерестовые участки имеют в длину и
ширину десятки метров и размеры нерестовых гнезд не ограничены.
По данным СВ. Красикова (1993), в относительно небольших притоках
бассейна Мезени средняя ширина нерестовых участков лосося определяется
размерами порогов и составляет 6 м при колебаниях от 3 до 5 м, длина
2-5 м, глубины варьируют от 0,4 до 1,0 м, грунт представлен мелкой и
средней галькой 2 - 4 см в поперечнике. В этих условиях нерестовые гнезда
формируются на относительно крутых склонах русла при переходе от
плеса к порогам (рис. 68). Ограниченные площади нерестовых участков и
мелкий нерестовый грунт обусловливают четкое подразделение бугров на
нерестовую яму, длина которой соответствует длине нерестящейся самки, и
собственно нерестовый бугор, высота которого может достигать 40 см.
Перекапывание ранее построенных нерестовых гнезд наблюдается в
единичных случаях (Владимирская, 1957). Даже при закладке икры на
ограниченных по площади нерестовых участках малых рек бассейна Мезени
самки лосося не разрушают бугор, построенный другими особями
своего вида (Красиков, 1993). Бугры близко расположенных друг к другу
гнезд могут соприкасаться или частично перекрываться краями, но нерестовые
ямы не накладываются друг на друга (см. рис. 68). При проведе-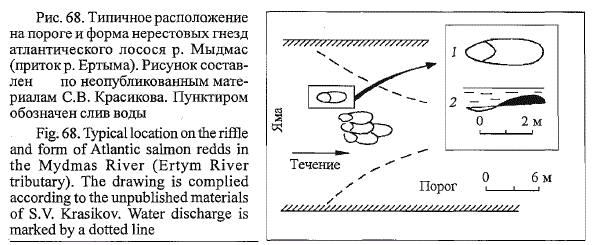
Рис. 68. Типичное расположение
на пороге и форма нерестовых гнезд
атлантического лосося р. Мыдмас
(приток р. Ертыма). Рисунок составлен
по неопубликованным материалам
СВ. Красикова. Пунктиром
обозначен слив воды
Fig. 68. Typical location on the riffle
and form of Atlantic salmon redds in
the Mydmas River (Ertym River
tributary). The drawing is complied
according to the unpublished materials
of S.V. Krasikov. Water discharge is
marked by a dotted line
нии исследований р. Щутор мы также не отмечали случаев перекопки
нерестовых бугров.
Посленерестовый скат производителей печорского лосося начинается
сразу же по окончании нереста, и часть вальчаков достигает предус-
тьевых районов моря той же осенью. В Коровинской губе 6-17 декабря
1974 г. были выловлены четыре самца-вальчака, которые отнерестились
осенью того же года в бассейне Печоры, поскольку других лососевых рек
поблизости нет (Антонова, Мартынов, 2004). Посленерестовый скат вальчаков
приостанавливается с наступлением ледостава, о чем косвенно свидетельствуют
случаи крайне редкого вылова вальчаков в осенне-зимнее
время в годы (до 1972 г.), когда на Печоре существовал промышленный и
любительский лов скатывавшихся после нереста сиговых рыб. Часть вальчаков
проводит зиму в таких удаленных от моря нерестовых реках, как
Щутор (наши данные) и Илыч (опросные данные). Так, в середине июня
1974 г. три вальчака (две самки и один самец) были выловлены в р. Щу-
гор в 40 км от устья. Все рыбы имели тусклую коричневую окраску тела,
белесовато-розового цвета мышцы и погруженную в кожные покровы
чешую. Их гонады находились на III - VI стадиях развития. В полости
тела самок имелась хорошо сохранившаяся без признаков резорбции и
жирового перерождения остаточная икра.
Массовый скат вальчаков, видимо, приходится на время весеннего
половодья. По опросным данным, со спадом уровня воды в весенне-летний
период они лишь единично встречаются в промысловых уловах в
магистральном русле Печоры. Отдельные вальчаки могут задерживаться
в реке на длительное время (Мартынов, 1983). Так, в сентябре 1974 г. в
р. Щутор в 45 км от устья была выловлена самка-вальчак, которая не могла
принадлежать к вальчакам этого года нереста, так как он начался только
18 сентября. Данная самка отличалась от лошалых рыб голубовато-
серебристой окраской тела и слабо погруженной в кожные покровы чешуей,
которая была сильно разрушена по переднему краю и не имела пос-
ленерестового прироста. Яичники находились на III - VI стадиях зрелос-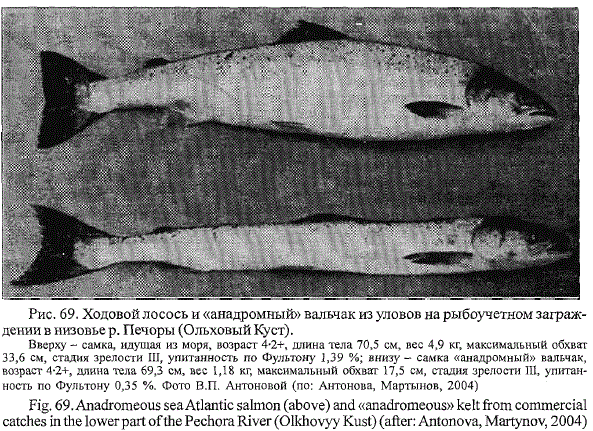
Рис. 69. Ходовой лосось и «анадромный» вальчак из уловов на рыбоучетном заграждении
в низовье р. Печоры (Ольховый Куст).
Вверху - самка, идущая из моря, возраст 4-2+, длина тела 70,5 см, вес 4,9 кг, максимальный обхват
33,6 см, стадия зрелости III, упитанность по Фультону 1,39 %; внизу - самка «анадромный» вальчак,
возраст 4-2+, длина тела 69,3 см, вес 1,18 кг, максимальный обхват 17,5 см, стадия зрелости III, упитанность
по Фультону 0,35 %. Фото В.П. Антоновой (по: Антонова, Мартынов, 2004)
Fig. 69. Anadromeous sea Atlantic salmon (above) and «anadromeous» kelt from commercial
catches in the lower part of the Pechora River (Olkhovyy Kust) (after: Antonova, Martynov, 2004)
ти. Коэффициент зрелости составил 0,52 упитанности по Фультону - 0,55.
Мышцы рыбы были белесоватого цвета. В полости тела и яйценосных
пластинках обнаружено 20 крупных остаточных икринок прошлогоднего
нереста, которые находились на стадии жирового перерождения, имели
неправильную угловатую форму и белесый цвет. Гонады с икрой новой
генерации были обильно пронизаны кровеносными сосудами и имели
темно-красный цвет. Состояние гонад свидетельствует о длительном
пребывания самки после нереста в реке. Если в 1973 г. нерест дгуторского
лосося закончился в конце сентября, то данная рыба провела после нереста
в реке 11-12 мес. Анализ структуры чешуи показал, что она нерестилась
дважды. Рыбы, подобные описанной выше, у местного населения
получили название «бывалец».
В низовьях Печоры отмечены единичные случаи захода в ловушки
РУЗа вместе с анадромными мигрантами серебристых вальчаков (Антонова,
Мартынов, 2004). «Анадромные» вальчаки отличались от нерестовых
мигрантов чрезвычайной истощенностью (рис. 69). Все они были
повторно отнерестившимися самками. Вовлечение отдельных вальчаков
в нерестовый ход лосося следует рассматривать как проявление стайного
поведения рыб.
В р. Печенге скат вальчаков весной через рыбоучетное заграждение за
5 лет наблюдений отмечен с 11 июня по 22 июля. В р. Варзуге, как пока-
168
зали результаты мечения (Мельникова, 1962; Бакштанский, Яковенко*
1976), он начинается вскоре после ледохода и приходится на вторую по-
ловинумая-июнь. По данным тех же авторов, случаи поимки вальчаков
осенью очень редки.
В р. Сидоровке отнерестившиеся производители скатываются в море
в июне, и пик их миграции предшествует началу массового хода лосося на
нерест (Кузьмин, 1985). Вальчаки туломского лосося в большом количестве
появляются в верхнем бьефе плотины Нижнетуломской ГЭС в начале
июня одновременно с покатной молодью и встречаются здесь в течение
всего лета до глубокой осени. Задержка части вальчаков перед гидросооружениями
Нижнетуломской ГЭС и, возможно^ их травмирование при
прохождении турбин могли способствовать повышению посленересто-
вой смертности и снижению доли остатка в популяции (см. гл. 3).
Смотрите также