ОБРАТНОЕ РАСЧИСЛЕНИЕ ЛИНЕЙНОГО РОСТА АТЛАНТИЧЕСКОГО ЛОСОСЯ ПО ЧЕШУЕ
В отличие от прямых натурных наблюдений, дающих представление
о групповом росте рыб, ретроспективная оценка позволяет получить
информацию об индивидуальном росте, что открывает дополнительные
возможности для анализа биологических закономерностей.
Например, изучение компенсационного роста рыб, анализ соотношения
речного и морского роста проходных рыб не представляются возможными
или весьма затруднительны без применения методов обратного
расчисления роста.
Обратному расчленению роста рыб посвящена обширная отечественная
и зарубежная литература, подробный обзор которой приводится в
ряде работ (Брюзгин, 1969; Мина, Клевезаль, 1976; Ваганов, 1978). Наиболее
перспективным считается метод номограмм, основанный на математическом
описании эмпирических связей между линейным ростом
рыбы и ее чешуи (Шентякова, 1966; Брюзгин, 1969; Малкин, 1999).
При ретроспективной оценке роста атлантического лосося ряд исследователей
пользуется формулой прямой пропорциональности Э. Леа, считая,
что она удовлетворительно отражает зависимость между ростом тела
и чешуи (Лишев, Римш, 1961; Евтюхова, 1970). Для балтийского лосося из
рек Швеции показано (Lindroth, 1963), что при переходе из реки в море
рост рыб ускоряется по сравнению с ростом чешуи, но сохраняет прямолинейную
зависимость. При этом возрастает угловой коэффициент прямой,
отражающий связь «тело-чешуя» в морской период жизни. В результате
расчисление роста по формуле Э. Леа приводит к завышению расчетной
длины покатника по сравнению с действительной. А. Линдрот
(Lindroth, 1963) предложил для приведения расчисленных по формуле
прямой пропорциональности длин смолта к реальным ряд переходных
коэффициентов со значениями 0,83 - 0,87 в зависимости от расчисленных
длин смолта (чем больше расчисленная длина смолта, тем меньше
коэффициент). Расчисление роста за речной период жизни осуществлялось
им по формуле Розы Ли:
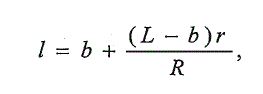 (1)
(1)где / - искомая длина пестрятки; L - длина смолта; г - радиус чешуи и
пестрятки; R — радиус чешуи смолта; Ъ— длина рыбы при закладке чешуи,
равная 18 мм (поправка Розы Ли).
По данным Д.К. Халтурина (1967), расчисленные методом Линдрота
(Lindroth, 1963) длины ладожского лосося (Salmo salar morpha sebago Girard)
хорошо согласуются с результатами прямых измерений.
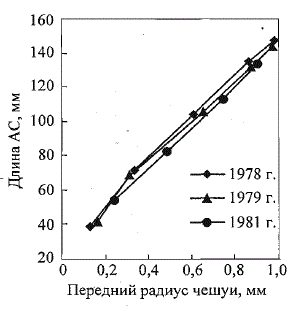
Рис. 14. Эмпирические кривые соотношения
«тело-чешуя» у молоди атлантического лосося
р. Щугор в разные по гидрологическим условиям
годы (по: Мартынов, 1983)
Fig. 14. Empirical curves reflecting the bodyscale
relationship in Atlantic salmon parr from the
Shugor River, caught in the years differing by
hydrological conditions (after: Martynov, 1983)
Для молоди лосося из рек Канады
показана криволинейная связь между
ростом тела и чешуи: L = 24 + 129г +
+ 122^(1, и г измерены в мм; Kerr, 1961).
Для молоди лосося из р. Шугор (бассейн
Печоры) выявлена аналогичная зависимость (Мартынов, 1983), причем
ее количественные параметры меняются в различные по гидрологическим
условиям годы (рис. 14). Приведенный выше слабо выраженный криволинейный
характер связи между линейным ростом тела и чешуи атлантического
лосося в речной период жизни во многом связан с феноменом
Розы Ли, а именно с затухающим по мере увеличения размеров рыб
влиянием длины малька при закладке чешуи. Если соотносить с длиной
переднего радиуса чешуи длину рыб за вычетом длины малька при закладке
чешуи, то показатель отношения длины тела к переднему радиусу
чешуи выравнивается у рыб разного возраста, и его связь с длиной тела
становится малозначимой (рис. 15).
Прямолинейный характер связи между длиной тела и чешуи сохраняется
в морской период жизни. При этом у рыб происходит увеличение
углового коэффициента прямой, отражающей связь «тело - чешуя» (рис.
16). Как показано нами ранее (Мартынов, 1983), поправочные коэффициенты
к расчисленной по чешуе производителей длине смолта, применявшиеся
Линдротом, имеют популяционную специфику и не могут быть
использованы для обратного расчисления длины смолта печорского лосося
по чешуе производителей. Наши данные по другим рекам свидетельствуют
о существенных межпопуляционных различиях угловых коэффициентов
уравнений первого порядка, отражающих связь «тело-чешуя» у
атлантического лосося (см. рис. 15). В связи с этим для оценки длины
молоди по чешуе производителей возможно использование коэффициента,
отражающего отношение длины тела за вычетом длины малька при
закладке чешуи молоди к переднему радиусу ее чешуи ((АС - b)/R). Данный
показатель во многом утрачивает популяционную и возрастную специфику
и становится близким к 125 (табл. 14).
Таким образом, межпопуляционная изменчивость отношения длины
тела к переднему радиусу чешуи (AC/R) определяется межпопуляцион-
ной изменчивостью длины малька при закладке чешуи и может быть
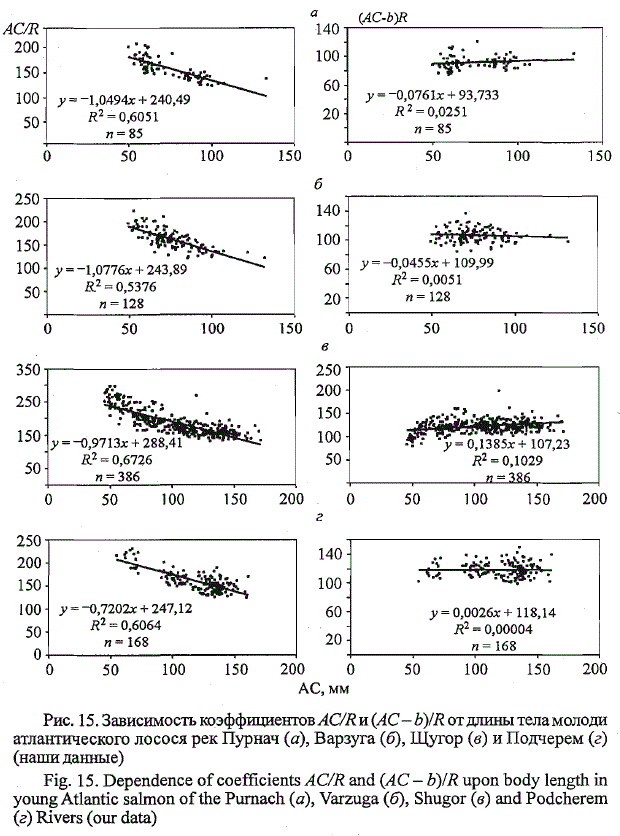
Рис. 15. Зависимость коэффициентов AC/R и (AC- b)/R от длины тела молоди
атлантического лосося рек Пурнач (а), Варзуга (б), Щугор (в) и Подчерем (г)
(наши данные)
Fig. 15. Dependence of coefficients AC/R and (AC - b)IR upon body length in
young Atlantic salmon of the Purnach (a), Varzuga (6), Shugor (e) and Podcherem
(г) Rivers (our data)
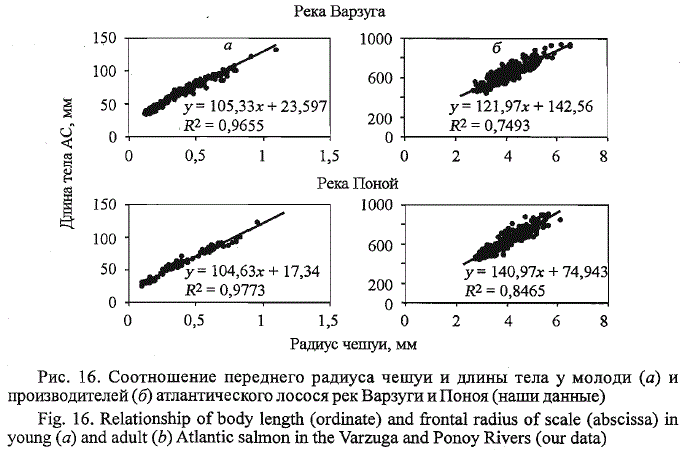
Рис. 16. Соотношение переднего радиуса чешуи и длины тела у молоди (а) и
производителей (б) атлантического лосося рек Варзуги иПоноя (наши данные)
Fig. 16. Relationship of body length (ordinate) and frontal radius of scale (abscissa) in
young (a) and adult (b) Atlantic salmon in the Varzuga and Ponoy Rivers (our data)
сведена к минимуму, если учитывать поправку Розы Ли. В этом случае
формула для оценки длин молоди по чешуе взрослых рыб, включая длину
смолта, будет иметь следующий вид: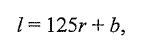 (2).
(2).
где / - длина молоди; r - длина переднего радиуса чешуи молоди; b- поправка
Розы Ли, имеющая популяционную (региональную) специфику.
Приведенное выше уравнение может быть использовано для расчис-
ления роста в речной период жизни по чешуе производителей лишь при
условии взятия чешуи из определительного участка, где изменчивость
размеров чешуи относительно невысока.
Обратное расчисление роста в морской период жизни может быть
удовлетворительно выполнено по формуле прямой пропорциональности
Э. Леа (цит по: Lindroth, 1963):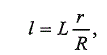 (3)
(3)
где / - искомая длина рыбы в морской период жизни; L - длина взрослой
рыбы при поимке; r - радиус чешуи, соответствующий искомой длине;
R - радиус чешуи рыбы при поимке.
Если при использовании чешуи взрослых рыб формула Э. Леа дает
удовлетворительную ретроспективу линейного роста лосося за морской
49
Таблица 14. Отношение длины тела к переднему радиусу чешуи у молоди атлантического
лосося на Севере России (наши данные)
Table 14. The dependence of body length - frontal radius of scales in young Atlantic salmon in
North of Russia (our data)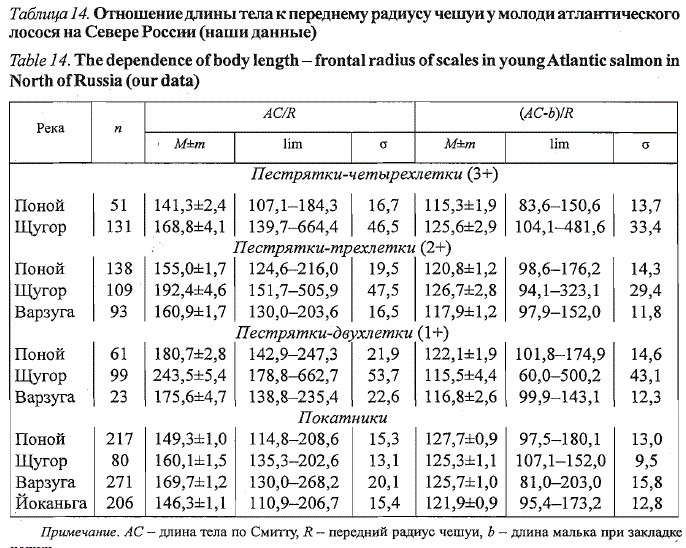
Примечание. АС - длина тела по Смитгу, R — передний радиус чешуи, Ъ — длина малька при закладке
чешуи.
период жизни, то при оценке роста за речной период жизни по формуле
(2) необходимо знать длину малька при закладке чешуи. Межпопуляци-
онная изменчивость поправки Розы Ли, рассмотренная выше, предполагает
популяционный подход к обратному расчислению роста атлантического
лосося. К сожалению, мы не располагаем прямыми данными по
длине малька при закладке чешуи для лосося рек Умбы, Йоканьги, Колы,
Туломы, Уры, Западной Лицы, Печенги, среднегодовые показатели теплосодержания
воды в которых изменяются от 1338 в р. Ура до 1702 гра-
дусо-дней в р. Туломе (см. Приложение, табл. 1). Мы сочли возможным
использовать при ретроспективной оценке линейного роста лосося этих
рек вычисленные показатели значений длины малька при закладке чешуи,
которые получены при подстановке в уравнение линейной регрессии
(см. рис. 11) суммы градусо-дней тепла воды в соответствующей реке
из табл. 1 Приложения.
При обратном расчислении роста печорского лосося, выловленного в
устье реки, использован усредненный показатель значений поправки Розы
Ли (28,4 мм), полученных при исследовании молоди в верховьях самой
Печоры и ее притоках: Подчерем, Щугор, Пижма. В результате при рас-
числении линейного роста лосося в реках Севера России были приняты
следующие значения длин мальков при закладке чешуи:
Река Коэффициент b, мм Река Коэффициент b, мм
Печора 28,4 Умба 25,4
Щугор 30,9 Йоканьга 28,8
Подчерем 28,9 Кола 26,5
Пижма 27,5 Тулома 24,1
Варзуга 24,7 Ура 29,1
Поной 26,9 Зап. Лица 26,4
Пурнач 26,9 Печенга 27,9
По формуле (2) с использованием приведенных выше поправок Розы
Ли определяли длину смолта (покатника) и пестряток. Рост рыб за морской
период жизни расчислялся по формуле прямой пропорциональности
Э. Леа (3), а по чешуе молоди - по формуле (1). При этом обозначению
«L» в формуле соответствуют значения длин молоди при поимке.
На примере молоди печорского лосося нами показано, что при расчете
длин пестряток с использованием формулы прямой пропорциональности
по чешуе из различных участков тела полученные значения средних
показателей длин достоверно не отличались (Мартынов В., Мартынов
Н., 1979). Данное обстоятельство свидетельствует о том, что отдельные
чешуйки на теле рыб при различиях в скорости роста увеличиваются
в размерах пропорционально росту длины тела. Поэтому при использовании
для расчисления роста рыб чешуи неизвестной топографии (коллекционные
материалы прошлых лет) целесообразно вести расчисление
по формулам прямой пропорциональности Э. Леа и Розы Ли. Однако при
этом следует помнить, что в результате будут получены несколько завышенные
показатели длин смолта, с одной стороны, и заниженные длины
годовиков - с другой.
Смотрите также