ОСНОВНЫЕ ФАКТОРЫ ЕСТЕСТВЕННОГО ВОСПРОИЗВОДСТВА АТЛАНТИЧЕСКОГО ЛОСОСЯ SALMO SALAR L. В БЕЛОМОРСКИХ РЕКАХ КОЛЬСКОГО ПОЛУОСТРОВА
Развитие Арктической зоны РФ оказывает существенное влияние на биологиче-ские ресурсы этого региона. Важной частью этих ресурсов являются анадромные рыбы и, прежде всего, атлантический лосось. Сохранение и эксплуатация популяций этого вида требует знания основных факторов, определяющих его естественное воспроизвод-ство.
Значительная часть ареала атлантического лосося расположена на территории Мурманской области и в Карелии, где этот вид воспроизводится в реках бассейнов Ба-ренцева и Белого морей, а также озер –Ладожского, Онежского, Куйто, Каменное и Се-гозеро. Первые обобщения по распространению лосося на Северо-Западе России были сделаны еще в XIX веке (Озерецковский, 1812; Данилевский, 1862, 1875; Кесслер, 1864, 1868; и др.). Затем в первой половине XX века наиболее крупный вклад в исследование лосося внесли академик Л.С. Берг (1916, 1923, 1932, 1935, 1948), известные ихтиологи В.Р. Аллеев (1913, 1928) и В.К. Солдатов (1920). Тогда же с 30-х годов начались систе-матические исследования биологии лосося в морской и речной периоды жизни (Труды КНИРС, 1935; Новиков, 1936; 1947; 1953; Казаков, 1982; Казаков, Веселов, 1998; Чер-ницкий, 1993; Калюжин, 2003; Мартынов, 2007 и др.). Однако в этих работах недоста-точно были освещены особенности воспроизводства лосося и характеристики нересто-во-выростных участков (НВУ) в различных по гидрологии речных и озерно-речных си-стемах. Инвентаризация лососевых рек и создание баз данных по факторам естествен-ного воспроизводства этого вида практически не проводились. Учет биологических ре-сурсов в специализированных ГИС и базах данных обеспечивает основу мониторинга и прогноза состояния биоресурсов. Это перспективное направление развивается в ряде зарубежных исследовательских центров (Baltic Salmon Rivers, 1999; Der Lachs…, 2008). Оно было начато и в нашей стране (Казаков, 1983; Веселов, Калюжин, 2001; Биология, …., 2005; Веселов, 2006; Зубченко и др., 2007; Калюжин и др., 2009; и др.). Создание базы данных и ГИС «Лососевые реки и популяции рыб Европейского Севера России» является одной из задач нашего проекта «Экологические основы сохранения естествен-ного воспроизводства и биоразнообразия анадромных рыб арктической зоны РФ», вы-полняемого по программе «Поисковые фундаментальные научные исследования в ин-тересах развития Арктической зоны Российской Федерации».
Для выполнения поставленной цели был проведен многомерный статистический анализ количественных параметров указанной выше базы данных. Всего в базу вклю-чено 147 рек бассейнов Баренцева, Белого и Балтийского морей, Ладожского и Онеж-ского озёр. В данной работе для анализа были взяты только беломорские реки Кольско-го полуострова (рис. 1) — 24 реки, по которым имеются наиболее полные сведения.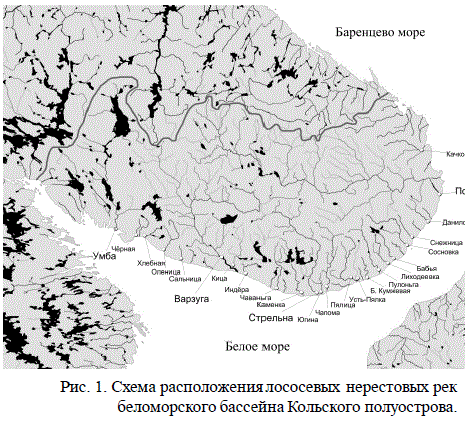
Рис. 1. Схема расположения лососевых нерестовых рек
беломорского бассейна Кольского полуострова.
Для анализа из указанной базы данных были выбраны 20 экологических показа-
телей, отражающих гидрологические и биологические характеристики лососевых рек:
длина реки (км), площадь водосбора реки (км2), падение (разница высоты русла реки
над уровнем моря в истоке и устье, м), относительное падение (м/км), общая озерность
бассейна реки (%); модуль стока (л/с/км2); среднегодовой расход воды в устье (м3/с),
площадь НВУ атлантического лосося (м2); число притоков I порядка, влияние рыбо-
ловства (экспертная оценка в баллах), лесистость бассейна реки (%), заболоченность
бассейна реки (%), фактическая численность производителей атлантического лосося
(экз.), фактическая численность смолтов лосося (экз.), средняя плотность молоди ат-
лантического лосося на НВУ (экз./100 м2), потенциальная численность смолтов (экз.);
потенциальная численность производителей (экз.), число видов рыб и возраст смолти-
фикации (год). Курсивом выделены расчетные показатели, остальные параметры были
получены в результате проведенных исследований.
Сбор материала для базы данных осуществляли в 23 экспедициях в период с
1999 по 2014 гг. Для сбора и обработки ихтиологических материалов использованы
стандартные и оригинальные методики (Правдин, 1966; Митанс, 1971, 1973; Смирнов,
Шустов, 1978; Павлов и др., 1999, 2007; Обзор методов…, 2000; Веселов, Калюжин,
2001). Основная часть этих методов изложена в руководстве, созданном с нашим уча-
стием (Кузищин и др., 2009). Численность производителей лососевых и процент их вы-
лова от численности нерестового стада определяли по результатам мечения, и по ре-
зультатам учета рыб на рыбоучетных заграждениях (РУЗ), по данным литературы (Зуб-
ченко и др., 1991; Калюжин, 2003; Реестр…, 2011), по отчетам рыбохозяйственных и
научных организаций о вылове рыб, по опросным и архивным материалам из фондов
ПИНРО и ФГУ «Мурманрыбвод». Плотность молоди лососевых и других видов рыб
оценивали методом электролова на контрольных НВУ разного типа (Zippin, 1958).
Маршрутные съемки рек осуществляли на лодках от истока до устья, комбинируя их с
пешим, автомобильным и вертолетным обследованием. Измеренные гидрологические
3
характеристики НВУ наносили на 3-D топографические схемы (Кузьмин, 1974, 1985; Шустов, 1983). Часть работ по обследованию НВУ выполняли с использованием видео-техники с дальнейшей расшифровкой данных. Оценивали площади НВУ и их качество: фракционный состав грунтов, скорости течения, глубины, степень заиления и зараста-ния (Обзор методов…, 2000). Потенциальную численность производителей и смолтов атлантического лосося вычисляли как произведение площади НВУ на эмпирический коэффициент, полученный на основании комплексной оценки (Power, 1973) ряда пока-зателей, включая данные о кормовой базе молоди лосося, плотности молоди разных возрастных групп.
Основные гидрологические характеристики рек были взяты из литературы (Гри-горьев, 1933; Ресурсы…, 1972; Каталог рек…, 1962; Берсонов, 1960) уточнены и до-полнены в ходе экспедиционных исследований.
Влияние отдельных факторов выявляли путем статистической обработки мате-риалов собранной базы данных стандартными методами корреляционного, кластерного и регрессионного анализа при помощи пакета программ Statistica 7. При проведении регрессионного анализа использовали метод «анализа сверху». Он заключается в том, что первоначально в анализ включали все количественные характеристики рек, а затем последовательно исключали наименее значимые параметры, до получения уравнения со всеми достоверными коэффициентами. Этот метод позволяет выявить все значимые факторы в анализируемой выборке.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
На основе проведенного анализа выявлено, что весьма велика вариабельность основных гидрологических параметров исследуемых рек и площади НВУ — последняя различается на три порядка; площадь водосбора, длина рек, расход воды — на два по-рядка. Биологические показатели различаются не меньше, чем гидрологические, например, по численности производителей популяции различаются в десятки тысяч раз. Это не позволяет без многомерных статистических методов выявить ведущие фак-торы исследуемых явлений. Поэтому в работе применены кластерный и многофактор-ный регрессионный анализы.
Систематизация лососевых рек.
Систематизация лососевых рек проводилась неоднократно, но она носила, в ос-новном, качественный характер и была основана на небольшом количестве классифи-цирующих признаков. Одна из наиболее известных классификаций была предложена С.В. Григорьевым (1933). Он, учитывая расположение озер и притоков, соединяющих их с главным руслом, выделил 3 типа речных систем. Первый тип – крупные озера и большая часть второстепенных озер расположены в верхней трети бассейна реки. Вто-рой тип – озера расположены почти равномерно или каскадом по системе реки. Третий тип – река служит стоком крупного озера. Однако реки характеризуются и другими гидрологическими показателями, имеющими значение для воспроизводства лосося. В связи с этим, нами (Веселов, 2006) в существующую классификацию был введен пока-затель «разветвленность», что позволило среди рек различной протяженности и гидро-логии выделить 7 основных типов: I. Малые неразветвленные или с 1–2 притоками без озер; II. Средние и крупные, сильно разветвленные, озерность менее 3%; III. Неразветв-ленный сток крупного озера; IV. Крупное озеро в устье; V. Разветвленный сток крупно-го озера; VI. Разветвленный сток группы озер,; VII. Разветвленный сток каскада озер.
Кластерный анализ показал, что беломорские реки Кольского полуострова раз-деляются на четыре одинаковых кластера, как по всем показателям базы данных, так и только по площади НВУ и гидрологическим характеристикам рек (рис. 2).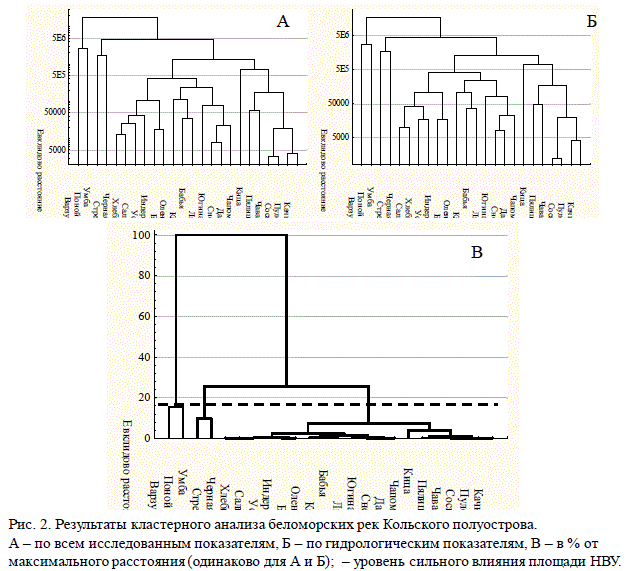
Рис. 2. Результаты кластерного анализа беломорских рек Кольского полуострова.
А – по всем исследованным показателям, Б – по гидрологическим показателям, В – в % от
максимального расстояния (одинаково для А и Б); – уровень сильного влияния площади НВУ.
На уровне 16% от максимального различия реки распадаются на два простых и
один смешанный кластер, который в свою очередь разделяется на два компактных кла-
стера на уровне 8%. Таким образом, беломорские лососевые реки разделяются на 4 кла-
стера. К первому относятся реки Поной и Варзуга; ко второму — Умба и Стрельна; к
третьему — Чапома, Пялица, Кица, Качковка, Пулоньга, Сосновка, Чаваньга; к четвер-
тому — Оленица, Бабья, Каменка (2), Лиходеевка, Даниловка, Снежница, Югина, Б.
Кумжевая, Индера, Сальница, Черная, Хлебная, Усть-Пялка. Следует отметить, что
уровень различий кластеров отражает степень сходства объектов классификации — чем
меньше этот уровень, тем более сходны объекты одного кластера и тем больше они от-
личаются от объектов других кластеров. Полученный уровень различий кластеров (16%
и 8%) является весьма хорошим для биологических исследований — чаще всего встре-
чаются различия на уровне 30–60% (Павлов и др., 2007). Наличие компактных класте-
ров с одной стороны указывает на то, что в анализе учтены действительно все основ-
ные факторы, влияющие на исследуемые явления. С другой стороны, оно дает возмож-
ность выделить реперные реки, на которых следует проводить мониторинг состояния
популяций. Выделение реперных объектов позволяет существенно сократить времен-
ные и финансовые затраты на мониторинг, без снижения необходимой точности.
Значение различных факторов для систематизации лососевых рек не одинаково.
Из исследованных 20 параметров только 7 показателей вносят достоверный вклад в
классификацию рек (табл. 1).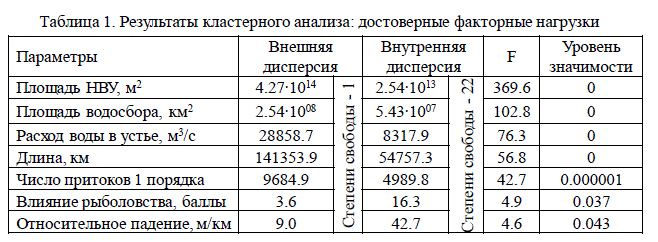
Таблица 1. Результаты кластерного анализа: достоверные факторные нагрузки
Такие факторы как падение реки, заболоченность и общая озерность, лесистость,
модуль стока и др. оказались статистически незначимы. Причем незначимость расчет-
ных параметров (например, модуль стока) обусловлена чисто математическими причи-
нами, а незначимость независимых параметров (заболоченность, озерность) имеет био-
логическое или гидрологическое содержание.
Учтенные в базе данных биологические показатели состояния популяций оказа-
лись не значимыми для количественной классификации. Причина этого заключатся в
том, что биологические показатели сильно связаны с многими другими показателями
— площадью НВУ и перечисленными в таблице 1 гидрологическими параметрами.
Рассмотрим подробнее значение основных (достоверных) факторов в порядке
уменьшения их факторных нагрузок.
Площадь НВУ. Анализ факторных нагрузок (табл. 1) показал, что наиболее
значимым параметром классификации является площадь НВУ. Этот показатель у рек
первого кластера на порядок больше, чем у рек второго кластера. Столь же велики раз-
личия между реками второго кластера и реками третьего и четвертого кластеров. У рек
третьего кластера площадь НВУ в 3,2 раза больше, чем у рек четвертого кластера.
Суммарно доступная для воспроизводства площадь нерестовых и выростных
участков беломорских рек Кольского полуострова составляет 53 млн. м2. Минимальная
площадь НВУ наблюдается в небольших реках, таких как Рязанка (93000 м2) и Усть-
Пялка (99000 м2). Наибольшими площадями НВУ обладают реки: Поной (17 340
000 м2), Варзуга (14 680 000 м2), Умба (4 478 000 м2), Стрельна (2 800 000 м2); Чапома
(1 395 000 м2). В остальных реках площади НВУ изменяются в пределах от 110000 до
700000 м2.
Площадь НВУ используется для расчета ряда важных биологических показате-
лей состояния популяций лососей, например, потенциальной численности молоди, по-
тенциального запаса производителей и др. В то же время определение площади НВУ
весьма трудоемкий процесс. И для многих рек он еще не выполнен. Поэтому, есть
необходимость в экспресс-оценке этого показателя. Как показал регрессионный анализ,
площадь НВУ зависит всего от трех гидрологических параметров рек — длины, расхо-
да воды и относительного падения. Эта зависимость аппроксимируется следующим
уравнением:
SНВУ = 13954 d – 44 d2 – 372212 op + 55749 op2 + 58185 Q – 2180 Q2
где
6
d – длина реки, км;
op – относительное падение, м/км;
Q – расход воды в устье, м3/с;
SНВУ – площадь НВУ, м2.
Уравнение достаточно полно отражает влияние гидрологических показателей рек на площадь НВУ — R2 = 0.9991, при р<< 0.00001. Все коэффициенты уравнения достоверны при р<0.0003. Это указывает на два обстоятельства: во-первых, на коррект-ный выбор параметров рек для базы данных, а во-вторых, на пригодность предложен-ного уравнения для экспресс-оценки площади НВУ во вновь исследуемых реках.
Типы НВУ. Выполненные нами бонитировочные работы на реках показывают, что количество НВУ и их качественные характеристики в притоках или в главном русле определяются макрорельефом бассейна водосбора, микрорельефом конкретных поро-гов, уклонами, т.е. факторами, вместе формирующими аллювиальные наносы грунтов, режимы скоростей течения и глубины. По условиям гидрологии были выделены груп-пы НВУ разной продуктивности, имеющие сходство по скоростям течения, уклонам, рельефам дна, составу грунта, площадям и др. показателям. Типичные пороги, харак-терны для всех зональных групп рек, но в меньшей степени для рек с повышенной озерностью. Они имеют значительные уклоны русла, выраженный рельеф, галечно-валунный грунт с отдельными глыбами. Их протяженность от 0,4 до 3–8 км, глубины 0,3–1,2 м, скорости течения 1,0–2,5 м/с. В тундровых реках глубины редко превышают 0,9 м. Нерестовые площади на типичных порогах занимают не более 5–12% площади. Распределение молоди лосося в таежных реках в основном прибрежное, мозаично–агрегированное. В тундровых реках распределение наблюдается по всей площади поро-га и оно также мозаично-агрегированное. На поворотах рек численность рыб по внут-реннему радиусу выше. Такие пороги заселены молодью на 40–60% площади, при плотности 22–76 экз./100 м2. Мелководные пороги, типичны для тундровых и таежно-тундровых рек, имеют протяженность 150–300 м, глубины 0,2–0,9 м, скорости 0,8–1,3 м/сек, сглаженный рельеф и галечно-валунное дно. Такие пороги обладают обшир-ными нерестовыми площадями (30–85%) и часто заканчиваются небольшими бочагами или плесовидными расширениями. На них агрегированные группы пестряток распреде-лены относительно равномерно, плотность 20–70 экз./100 м2. Перекаты, чаще встреча-ются в реках таежной группы (Умба, Оленица, Сальница, Варзуга и др.), расположены на мелководных плато с гребнями и локальными перепадами глубин от 0,2 до 0,8 м, при скоростях 0,4–1,0 м/сек. Отличаются мелкими фракциями грунта. Нерест возможен на 60–90% площади. Площадь заселения молодью составляет 20–30%, чаще всего 5–10%. Обычно пестрятки концентрируются на гребнях (25–80 экз./100 м2). Малые при-токи (ручьи): расширяют территорию обитания молоди лосося. Характеризуются ве-сенним заселением всех возрастных групп молоди с других НВУ и осенней миграцией обратно в главное русло. Характеристики НВУ ручьев часто соответствуют лучшим показателям (если они впадают в порожистые участки рек), поэтому плотности молоди на них достигают 150-190 экз.100/м2. Однако протяженность заселяемых участков не превышает 200–300 м от устья. Нерест производителей в ручьях возможен в устье или на удалении от него до 300 м. Многочисленные малые притоки отмечаются в реках Варзуга, Поной и Умба. Следующие два типа НВУ отмелевые косы и протоки харак-терны как для крупных, так и небольших таежных рек, имеющих после порогов рав-нинные участки с небольшими уклонами (0.9–1.2 м/км). Это реки: Варзуга, Кица, Оле-ница, Сальница, Кузрека, Умба и др. Они, как правило, заселяются молодью лосося с других участков в весенний период. Распределение молоди на них крайне неравномер-ное и плотности от сезона к сезону широко варьируют. С падением уровня воды к сере-
дине лета сеголетки и старшие возрастные группы пестряток мигрируют на другие НВУ в главное русло. Нерест производителей возможен на напорном склоне отмелевых кос, а в протоках на валообразном подъеме дна, образующемся в местах сопряжения с главным руслом.
Анализ данных по распределению НВУ в руслах рек и сроков нерестового хода лосося показал, что для крупных разветвленных рек из первого кластера (Поной, Варзуга) и средних разветвленных речных систем из второго кластера (Умба, Стрельна) характерен осенний, весенний и летний тип нерестовых миграций, что согласуется с протяженностью рек и разной удаленностью нерестилищ от устья. В более коротких реках третьего кластера наблюдается преимущественно весеннее-летний ход, а в чет-вертом кластере – только весенний.
Площадь водосбора. Суммарно площадь водосбора исследуемых рек составля-ет 49764 км2. Наибольшая величина показателя принадлежит двум рекам из первого кластера (25303 км2), затем двум рекам из второго кластера (9022 км2), семи рекам из третьего кластера (7069 км2) и тринадцати рекам из четвертого кластера (3285 км2). Наименьший показатель принадлежит реке Рязанка (101,6 км2), а наибольший, в убы-вающей последовательности, рекам Варзуга (9836 км2) и Умба (6248 км2). Для осталь-ных рек площадь водосбора колеблется в пределах 100-1300 км2.
Среднегодовой расход воды в устье реки. Важная характеристика, указываю-щая на водность речной системы, что, в конечном счете, определяет разнообразие условия для воспроизводства лососевых рыб. Например, реки или непересыхающие ру-чьи с расходом воды менее 1,5 м/с населены только кумжей. Реки, с расходом воды от 1,5 до 3 м3/с, населены как лососем, так и жилой и проходной кумжей. Водотоки с рас-ходом более 3,5 м3/с считаются приоритетно лососевыми, а кумжа в таких водоемах воспроизводится преимущественно в притоках. По всем исследуемым рекам среднего-довой расход в устье составляет 24.4 м3/с. Среднее значение расхода максимально в первом кластере — 139 м3/с, во втором, третьем и четвертом кластерах он более чем в два раза ниже, соответственно, 55, 13 и 8 м3/с. Наибольший расход отмечен в реке По-ной (176.5 м3/с), а наименьший — в реке Большая Кумжевая (2.1 м3/с).
Длина реки. Общая протяженность беломорских рек Кольского полуострова 2559,1 км. Средние длины рек монотонно убывают от первого к четвертому кластеру и составляют 339,9, 169, 75,1 и 38,8 км соответственно. К наиболее крупным рекам отно-сятся: Поной (425,7 км), Варзуга (254 км), Стрельна (213,2 км), Умба (124,8 км) и Чапома (115,3 км). Их суммарная протяженность достигает 1133 км, или 44%. Протя-женность остальных речных систем суммарно составляет 1426,1 км и изменяется в пре-делах от 8,7 км (Колвица) до 91,6 км (Пялица), а в среднем составляет 43,2 км.
Число притоков I порядка существенно различается в реках выделенных кла-стеров. В реках первого кластера их в среднем 80, во втором кластере в 2.6 раза меньше — 31, в третьем кластере их число снижается еще в 3 с лишним раза — до 9, а меньше всего притоков в реках четвертого кластера — 2.
Классифицирующее значение числа притоков было отмечено и ранее. Так в ука-занной выше систематизации С.В. Григорьева (1933), модифицированной А.Е. Весело-вым (2006), классифицирующими признаками являются число и расположение озер и разветвленность речной системы (число притоков). Кластерный анализ, проведенный в данной работе, подтвердил значимость разветвленности (количества притоков), но при количественной оценке вклад озерности в систематизацию оказался незначимым. Это вполне объяснимо, так как воспроизводство (нерест и нагул молоди) атлантического лосося происходит в главном русле рек и притоках, а не в озерах.
Воздействие рыболовства уменьшается в порядке выделенных кластеров от пер-вого до четвертого. В среднем в реках первого кластера вылов достигает 3.5 балла
8
(60%), в реках второго кластера — 3 балла (до 50%), в третьем кластере — 2.4 балла (38%), а в четвертом —2 балла (27%). В наибольшей степени страдают от незаконного рыболовства запасы лосося в доступных для браконьеров реках.
Относительное падение в первом и втором кластерах почти одинаково (1.3 и 1.0 м/км), а в третьем и, особенно, в четвертом кластерах — значительно увеличивается (2.8 и 4.2 м/км). Для всех 4 кластеров беломорских рек Кольского полуострова в сред-нем абсолютное падение составляет 177.5 м, изменяясь в пределах от 159 до 236 м. Наименьшее падение у реки Пила (49 м), а наибольшее у реки Поной (292 м). Вместе с тем, река Поной имеет самый низкий показатель относительного падения – всего 0.69 м на 1 км длины реки. Поэтому, основное русло насыщено плесами и в меньшей степени порогами, пригодными для нереста лосося. В реке Поной нерестовое значение имеют в основном многочисленные притоки порогово-перекатной структуры и с относительным падением выше 1.5 м/км. Наибольшее относительное падение отмечено для реки Ор-ловка – 7.8 м/км длины, в которой структура русла представляет собой сплошной по-рог, с неглубокими бочагами и перекатами. В беломорских реках Кольского полуост-рова показатель относительного падения в среднем составляет 4.0 м3/с. В целом высо-кий показатель падения исследуемых рек свидетельствует о горном типе водотоков, характеризуемых наличием многочисленных порогов и перекатов, перемежающихся с плесовыми участками. Такая структура рек наилучшим образом отвечает требованиям лососевых нерестовых рек.
Анализ продольных профилей рек всех четырех кластеров показал, что для нереста лососем используется преимущественно участки, на которые приходятся по-следние перед устьем 60-80 м падения, за исключением некоторых протяженных рек с цепью озер (Умба), или в которых важны для воспроизводства лосося притоки I–II-ого порядков (Поной). У тундровых беломорских рек крутое падение формирует русло ре-ки практически без плесов и озеровидных расширений, что и определяет большие пло-щади НВУ и использование лососем до 70% протяженности главного русла.
Факторы, определяющие естественное воспроизводство атлантического лосося.
С целью выявления основных факторов определяющих естественное воспроиз-водство лосося были проанализированы регрессионные связи пяти биологических по-казателей (фактическая численность производителей и смолтов, возраст смолтифика-ции, средняя плотность молоди лосося на НВУ и число обитающих видов рыб) со все-ми остальными параметрами, учтенными в базе данных. Установлено, что достоверное влияние на биологические характеристики оказывают только те шесть факторов, кото-рые оказались значимыми для систематизации лососевых рек (табл. 1). В таблице 2 приведены уравнения, которые показывают, какие именно из этих шести факторов влияют на тот или оной биологический показатель.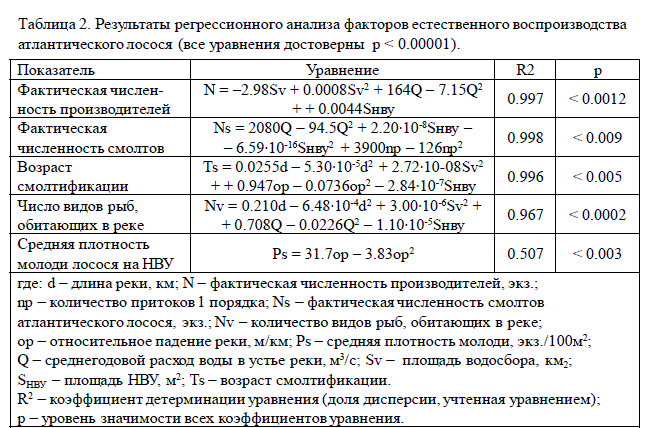
где: d – длина реки, км; N – фактическая численность производителей, экз.;
np – количество притоков 1 порядка; Ns – фактическая численность смолтов
атлантического лосося, экз.; Nv – количество видов рыб, обитающих в реке;
op – относительное падение реки, м/км; Ps – средняя плотность молоди, экз./100м2;
Q – среднегодовой расход воды в устье реки, м3/с; Sv – площадь водосбора, км2;
SНВУ – площадь НВУ, м2; Ts – возраст смолтификации.
R2 – коэффициент детерминации уравнения (доля дисперсии, учтенная уравнением);
р – уровень значимости всех коэффициентов уравнения.
Таблица 2. Результаты регрессионного анализа факторов естественного воспроизводства
атлантического лосося (все уравнения достоверны р < 0.00001).
Фактическая численность производителей атлантического лосося. В реках
первого кластера численность репродуктивной части популяций лосося составляет 30 –
55 тыс. экз., в реках второго кластера — 800 – 8500 экз., в реках третьего кластера она
колеблется от 1700 до 5500 экз, а в реках четвертого кластера она минимальна — 120 –
1500 экз.
Регрессионным анализом (табл. 2) установлено, что численность производите-
лей нелинейно зависят от площади водосбора реки, ее расхода и площади НВУ. Полу-
ченное уравнение имеет высокий коэффициент детерминации (R2), который показывает
долю учтенной дисперсии. Для приведенной в таблице 2 зависимости R2 = 0.997 то есть
она учитывает 99.7% от всех влияний на численность производителей. Это указывает
на высокую прогностическую силу уравнения и позволяет использовать его на практи-
ке.
Наиболее важны для воспроизводства атлантического лосося в беломорском
бассейне Кольского полуострова следующие реки (в порядке убывания численности
стада): Варзуга (55000 экз.), Поной (30000 экз.), Стрельна (8500 экз.), Сосновка
(3500 экз.), Качковка (2700 экз.), Пялица (2700 экз.) и Кица (2500 экз.). Рекордные чис-
ленности производителей на протяжении последних 70 лет отмечали в реке Варзуга
(75–135 тыс. экз.), а наименьшие – в малых реках Лувеньга (70 экз.), Рязанка (50 экз.),
Большая Кумжевая (120 экз.) и Орловка (50 экз.).
Фактическая численность смолтов атлантического лосося максимальное
значение имеет в реках первого кластера (565 250 экз. в среднем), уменьшается в реках
второго кластера до 106 300 экз. и на порядок меньше в реках третьем и четвертом кла-
стерах (52 800 и 12 454 экз. соответственно). Регрессионный анализ (табл. 2) показал,
что продукция смолтов также нелинейно связана с расходом реки, площадью НВУ и
числом притоков. Это уравнение имеет наивысший коэффициент детерминации, что
указывает на его высокую прогностическую ценность.
Возраст смолтификации может служить оценкой условий существования мо-лоди — чем больше этот возраст, тем хуже условия обитания в реке. Этот показатель слабо зависит от кластера, к которому принадлежит та или иная река. Он нелинейно зависит от длины реки, площади водосбора и профиля реки (относительного падения), а также от площади НВУ (табл. 2).
Число видов рыб, обитающих в реке. В беломорских реках Кольского полу-строва встречаются следующие виды рыбообразных и рыб: тихоокеанская минога Lampetra japonica, атлантический лосось Salmo salar, жилая и проходная формы кумжи Salmo trutta, ряпушка Coregonus albula, сиг Coregonus lavaretus, корюшка Osmerus eperlanus, хариус Thymallus thymallus, щука Esox lucius, плотва Rutilus rutilus, елец Leuciscus leuciscus, обыкновенный гольян Phoxinus phoxinus, налим Lota lota, колюшка девятииглая Pungitius pungitius, окунь Perca fluviatilis, ерш Acerina cernua, бычок четы-рехрогий Triglopsis quadricornis, камбала речная Pleuronectes flesus; камбала полярная Liopseta glacialis, арктический голец Salvelinus alpinus и интродуцированная горбуша Oncorhynchus gorbuscha.
В выделенных кластерах число сопутствующих видов изменяется не намного — его среднее значение монотонно уменьшается от первого к четвертому кластеру: 17.5, 15.0, 10.4, 8.8 видов. Регрессионный анализ (табл. 2) показал достоверную связь числа видов в реке с основными гидрологическими характеристиками рек: длиной, расходом воды, площадью водосбора и площадью НВУ. Полученная зависимость имеет хорошую прогностическую силу.
Рассматривая видовой состав рыб, следует отметить, в исследованных реках обитает от 4 до 20 видов. Однако на НВУ лосося обычно встречается не более 13 ви-дов, а чаше всего 3-6 видов. На выростных участках численность молоди лосося обыч-но составляет 85–95% по отношению к другим видам рыб.
Средняя плотность молоди атлантического лосося на НВУ в беломорских реках Кольского полуострова изменяется в пределах от 7 до 297 экз./100 м2. Наиболь-шая средняя плотность (89.3 экз./м3) отмечена на реках первого кластера, наименьшая (25.0 экз./м3) на реках второго кластера. На реках третьего и четвертого кластеров она составляет 60.9 и 67.5 экз./м3 соответственно.
Регрессионный анализ (по методу «анализа сверху») показал, что плотность мо-лоди связана только с относительным падением реки (табл. 2), при наименьшем значе-нии коэффициента детерминации полученного уравнения — им учтено только 50.7% от суммы влияния всех учтенных и неучтенных факторов. Наряду с этим корреляционный анализ показывает наличие связи (коэффициент корреляции Спирмена — 0.46, при р<0.05) плотности молоди с численностью производителей. Поэтому дополнительно был проведен отдельный регрессионный анализ связи этих показателей. Он дал следу-ющее уравнение: Ps = 0.0119N – 2.04∙10-07N2 (при R2 = 0.248), которое учитывает только 24.8% влияний от всех существующих. Низкая прогностическая сила полученных уравнений связана с тем, что плотность молоди складывается преимущественно из плотности сеголеток лосося. А она формируется в течение нескольких месяцев под воз-действием многих условий меняющихся от года к году. В результате плотность молоди атлантического лосося в большей степени зависит от лабильных характеристик экоси-стемы рек (определяемых условиями года), чем от параметров, учитываемых в базе данных. То есть, пользоваться полученными уравнениями для экспресс-оценки концен-трации молоди на новых реках пока невозможно.
В результате анализа биологических показателей, характеризующих состояние популяций атлантического лосося установлено, что такие показатели как фактические численность производителей и смолтов, возраст смолтификации и число обитающих
11
видов рыб сильно зависят от учтенных в базе данных гидрологических параметров рек, а плотность молоди на НВУ слабо с ними связана. Причина этого, на наш взгляд, в том, что четыре упомянутых показателя формируются в течение нескольких лет, от 3-4 лет (запасы смолтов), до нескольких десятилетий (число обитающих видов рыб). Поэтому лабильные условия разных лет усредняются и стабильные гидрологические показатели и площадь НВУ становятся определяющими для этих параметров. Плотность молоди на НВУ преимущественно формируется в течение текущего года, и поэтому она в большей степени зависит от лабильных условий года.
Интересно заметить, что площадь НВУ входит во все полученные уравнения, имеющие прогностическую силу. Кроме того, она вносит наибольший вклад в система-тизацию лососевых рек. Это указывает на то, что площадь НВУ является комплексным фактором, который интегрирует многие гидрологические и биологические факторы, определяющие условия существования лососевых популяций. НВУ определяет условия нереста и инкубации икры, масштабы расселения молоди, кормовую базу и условия нагула молоди, то есть основные условия естественного воспроизводства и формирова-ния численности поколений. Именно через количество и качество НВУ осуществляется опосредованное влияние гидрологических факторов (длина реки, расход воды, относи-тельное падение) на популяционные показатели атлантического лосося.
ЗАКЛЮЧЕНИЕ
В результате анализа количественных показателей из базы данных «Лососевые реки и популяции рыб Европейского Севера России» для 24 беломорских рек Кольско-го полуострова впервые была выполнена систематизация лососевых рек. На уровне 8-16% от максимального расстояния в пространстве анализируемых признаков были вы-явлены четыре компактных кластера объектов систематизации. Столь хороший для биологических исследований уровень различия кластеров дает основания для выделе-ния реперных (модельных) рек внутри каждого кластера. Это существенно упрощает организацию мониторинга состояния естественного воспроизводства атлантического лосося, сокращая время и финансовые затраты на его проведение.
Выявлена иерархия экологических факторов, определяющих состояние экоси-стем лососевых рек Арктической зоны. Из 20 исследованных только 7 оказались досто-верно значимыми факторами, определяющими типы лососевых рек (4 типа). Это такие показатели как площадь нерестово-выростных участков атлантического лосося (НВУ), основные гидрологические параметры его нерестовых рек (площадь водосбора, средне-годовой расход воды, длина, количество притоков 1 порядка и относительное падение русла рек), а также воздействие рыболовства. Биологические показатели сильно связа-ны с площадью НВУ и перечисленными гидрологическими параметрами. Они форми-руются в результате многолетних процессов роста и развития рыб. Такая длительность нивелирует лабильные сезонные и годовые изменения среды, что усиливает влияние постоянных гидрологических факторов.
Множественный регрессионный анализ подтвердил высокую значимость ука-занных параметров. Биологические показатели состояния популяций, которые форми-руются в течение трех лет и более (запас производителей и численность смолтов, воз-раст смолтификации, число видов рыб, обитающих в реке) определяются только зна-чимыми гидрологическими параметрами. При этом влияние этих параметров весьма высоко — 96.7-99.8% от силы всех влияний на исследуемый показатель. Выявленные зависимости позволяют оценивать пригодность рек для обитания и воспроизводства атлантического лосося, степень их использования этим видом, а также целенаправлен-но исследовать механизмы формирования численности его популяций.
Установлено, что площадь НВУ атлантического лосося является комплексным фактором, интегрирующим многие гидрологические и биологические факторы, определяющие условия существования лососевых популяций. От площади НВУ зависят все указанные выше показатели состояния популяций атлантического лосося. Определена регрессионная зависимость этого фактора от длины рек, расхода воды и рельефа русла (относительное падение). Данная зависимость имеет высокую прогностическую силу и может быть использована для экспресс-оценки площади НВУ этого вида во вновь ис-следуемых реках. Следует отметить, что данная работа выполнена только для части созданной ба-зы данных — для беломорских реках Кольского полуострова. В дальнейшем планиру-ется продолжить подобный анализ и для других частей ареала атлантического лосося. Работа выполнена при финансовой поддержке Программы фундаментальных исследований Президиума РАН «Поисковые фундаментальные научные исследования в интересах развития Арктической зоны Российской Федерации».
Д.С. Павлов1, А. Е. Веселов2, В.В. Костин1, Д.А. Ефремов2, М.А. Ручьев1
1Институт проблем экологии и эволюции им. А.Н. Северцова РАН
2Институт биологии Карельского научного центра РАН,
Этот адрес электронной почты защищён от спам-ботов. У вас должен быть включен JavaScript для просмотра.
Смотрите также