
РОСТ И ПОЛОВОЕ СОЗРЕВАНИЕ САМОК АТЛАНТИЧЕСКОГО ЛОСОСЯ SALMO SALAR L. БЕЗ НАГУЛА В ПРИРОДНЫХ ВОДОЁМАХ
Лососевые – факультативно голобиотические рыбы. Обычно они мигрируют в моря и, используя обширные кормовые ресурсы, достигают больших размеров, продуцируют крупную икру.
Если миграция невозможна, весь жизненный цикл осуществляется в пресной воде. Еще полтора столетия назад В. П. Врасский выращивал атлантического (балтийского и озерного) лосося до половой зрелости на Никольском рыбоводном заводе (р/з). Позднее аналогичные работы проведены в Шотландии, Дании, Швеции, Норвегии, США и других странах. Особенно интенсивно полноцикловое выращивание стало развиваться с конца 20-го века, когда для товарного лососеводства потребовались сотни миллионов штук икры, а заход производителей в реки сократился (Jansen, McLeary, 1996). В прибрежных водах Атлантики, характеризующихся круглогодично благоприятным гидрологическим режимом, маточные стада лосося обычно содержат в крупногабаритных морских садках и береговых бассейнах, но положительные результаты получают также в пресной воде на рыбоводных станциях Финляндии и Польши (Эскелинен, 1991а; Бартель и др., 2008). Успехи названных стран способствовали возрождению интереса к созданию жилых стад и в России.
Приобрели актуальность вопросы управления ростом и созреванием лосося на отечественных р/з, оценки биологической полноценности выращиваемых жилых особей в качестве замены проходным производителям при поддержании природных популяций. С этих позиций мы изучали пластичность репродуктивной функции в онтогенезе лосося (Мурза, 1979; Мурза, Оганесян, 1982; Мурза, Христофоров, 1982; Христофоров, Мурза, 1998а; и др.). Было показано, что гаметогенез в речной (заводской) и морской периоды жизни этого вида представляет собой единый процесс, не зависящий непосредственно от солёности воды. При задержке самок на рыбоводных предприятиях до достижения размеров, превышающих типичные для смолтов в период катадромной миграции, яичники у них продолжают развиваться, и наблюдается дифференциация особей на группы с разным темпом гаметогенеза. Созревающие самки пост-смолты лосося описаны нами в прудах Нарвского р/з (Христофоров, Мурза, 1985), а позднее развитие гонад у этого вида прослежено при опытно-производственном выращивании его до половой зрелости на других р/з ФГУ «Севзапрыбвод»: Лужском – с 1995 г., Невском – в 1998–2001 гг. и Свирском – в 1999–2005 гг.
Цель данного сообщения – сравнить биологические, в том числе репродуктивные, показатели жилых самок первого в Ленинградской области маточного стада лосося, созданного на Лужском производственно-экспериментальном лососевом рыбоводном заводе (ЛПЭЛРЗ), с таковыми проходных самок из популяций рек Невы и Нарвы, послуживших донорами при его формировании. Кроме того, 15 лет, прошедшие с начала формирования этого стада (Мурза и др., 1997; Мурза, Христофоров, 2000, 2004а)*, – достаточный срок для оценки его вклада в поддержание лужской популяции лосося. Предпосылки для создания маточного стада лосося на ЛПЭЛРЗ. В прошлом р. Луга играла важную роль в воспроизводстве и промысле лососевых. К началу 20-го века в ней и в Лужской губе, добывали ещё до 10 тыс. шт., а в 1930-е гг. – 3,0–3,5 тыс. шт. лосося (Гримм, 1905; Кучина, 1939; и др.). Сокращение численности популяции, как отмечалось (Христофоров, Мурза, 1998б, 2000а, б; 2002, 2003), произошло вследствие лесосплава, загрязнения вод промышленными стоками и наличия плотины кожевенно-картонажной фабрики «Партизан» у Кингисеппа, многие годы преграждавшей доступ производителям к основным нерестилищам. С 1930-х до начала 1950-х гг. обсуждалась необходимость строительства рыбохода и р/з. В 1947 г. на выбранной для завода площадке у впадения р. Хревицы в р. Лугу были заложены шурфы, но проектирование началось только в 1974 г., а строительство – в 1981 г. Газета «Ленинградская правда» сообщала 12 сентября 1981 г., что крупнейший в Ленинградской области р/з «... будет выпускать до полумиллиона двухгодовалой молоди лосося ежегодно. По проекту рыбоводный комплекс включает в себя цехи: инкубационный, выростной, приготовления кормов, лабораторию и другие подразделения. Всё это сложное хозяйство станут обслуживать всего 80 человек, так как большинство процессов предусматривает полную механизацию и автоматизацию». Ко времени пуска р/з в эксплуатацию 30 декабря 1989 г., производство уже морально устарело, а нативная популяция лосося р. Луги, с целью поддержания которой его строили за счёт средств компенсации ущерба от ЛАЭС, перестала существовать. Последние случаи поимки лужского лосося датированы 1964 и 1969 гг. К началу 1990-х гг. удавалось отлавливать только кумжу. Икру её привозили на ЛПЭЛРЗ также с рек Систы и Воронки. Для воспроизводства лосося сеголеток доставляли с Нарвского р/з, а икру – с рек Даугавы, Нарвы, Невы и из Финляндии. Проектную мощность р/з, предполагавшую ежегодный выпуск в р. Лугу и близлежащие реки 500 тыс. шт. двухгодовиков лосося и кумжи, снизили до 300 тыс. шт.
За счёт выпусков пестряток в возрасте 1÷1+ и 0+ началось формирование новых популяций, и с 1993 г. возобновился заход лосося в р. Лугу. В 1993–1994 гг. было учтено 27 и 3 шт. В 1995–1996 гг., после организации перекрытия у дер. Струппово, поймали 46 и 34 шт. Численность и возрастной состав производителей обнаруживали чёткую связь с динамикой выпусков молоди (Христофоров, Мурза, 2003). По данным анализа структуры склеритов речной зоны чешуи в разные годы, все лососи и от 54 до 70% особей кумжи имели заводское происхождение. Доля рыб с «нерестовыми марками» на чешуе постепенно увеличилась с 4,0 до 16,0 – 38,5%. Общая численность заходивших в р. Лугу производителей лосося достигла 50–100 шт., кумжи – 200–300 шт.
Для дальнейшего увеличения репродуктивного потенциала популяций обоих видов и обеспечения ЛПЭЛРЗ икрой требовался переход к более эффективным выпускам продукции на стадии смолта*. Он не был осуществлён, так как совещание российских и финских специалистов летом 1994 г. в Санкт-Петербурге изменило направление деятельности ЛПЭЛРЗ. Решено было использовать более чем 15-летний опыт Финляндии в эксплуатации маточных стад лосося для развития этого направления в нашей стране.
Приоритетным признали сохранение генофонда лосося р. Невы, но так как на Невском р/з условия для содержания стада отсутствовали, то избрали ЛПЭЛРЗ – самый новый и мощный в регионе. Завод располагал возможностью замены принудительной водоподачи на самотёчную. На нём предусматривались также подогрев воды для инкубации икры и подращивания личинок, система оборотного водоснабжения и охлаждения воды в цеху длительного выдерживания производителей (ДВП). Рыбоводно-биологическое обоснование (РБО) для формирования маточного стада подготовили в 1994 г. сотрудники ГосНИОРХ и АО «Форкис», взяв за основу «финскую» биотехнологию. Согласно сформулированным в РБО задачам, ЛПЭЛРЗ должен был стать центром, снабжающим икрой остальные р/з области, увеличить их мощность для обеспечения квот на вылов лосося в Балтийском море, снабдить посадочным материалом природоохранные мероприятия, связанные с восстановлением популяций в реках, и создать постоянный резерв такого материала для продажи, в том числе за рубеж. От стада численностью 1400 особей, включающего 7 генераций рыб (в возрасте от 0+ до 6+) и ежегодно пополняемого за счёт икры проходных самок, рассчитывали получать 1,4 млн. шт. икры лосося. Общий вес рыб стада должен был составить 4,176 тонн, потребность в воде – 4,09 м3/мин., в выростных площадях – 1725 м2. В том числе, требовались 16 бассейнов по 100 м2, отсутствовавшие на ЛПЭЛРЗ.
Формирование генераций и интенсификационные приемы. Маточное стадо предполагалось создать за счёт гамет лосося р. Невы, но фактически использовался и другой донорский материал.
_____________
* Исходя из размеров смолтов, расчисленных нами по чешуе возвращавшихся в р. Лугу производителей лосося, следовало выпускать весной молодь с длиной тела не менее 15,0 см, соответствующей стандарту, принятому в скандинавских странах.
1-я генерация стада (1993 г.) получена из икры лосося р. Луги. Для выращивания крупной молоди использовали подогрев воды. Средняя температура её при инкубации составляла 3,6 °С в январе и 2,1 °С в феврале 1994 г. Личинки вылупились в феврале – марте при 8,2 °С, тогда как в естественных температурных условиях это происходит в апреле. Кормить их начали во второй половине апреля. К началу июня рыб перевели в садки, а в конце октября вернули в основной цех на зимовку. Впервые в условиях ЛПЭЛРЗ были получены сеголетки лосося со средним весом 8,9 г, то есть в несколько раз больше нормативного (по «Инструкции ..., 1979» – 2,2–3,2 г). В конце зимовки воду подогревали до 11,0–12,0 °C, и в марте годовики стали серебриться. С учётом соотношения полов и численности карликовых самцов среди одновозрастной молоди разных размерных категорий, для стада в апреле 1995 г. отобрали 498 шт. средних по весу рыб (20,0 г).
2-я генерация (1994 г.) получена из икры лосося р. Невы. Её привезли на стадии «глазка». Личинки вылупились в апреле 1995 г. При работе с ними использовали подогрев воды. В конце мая молодь перенесли из лотков в бассейны, а с середины июля до начала ноября содержали в садках. После возвращения в цех сеголеток рассортировали. Для стада в апреле 1996 г. отобрали 1064 шт. средних по весу годовиков (21,5 г). Их перевезли в цех в Ивановском, где рыбы посеребрились.
3-я генерация (1995 г.), как и 1-я, исходно формировалась из икры лосося р. Луги. Подогрев при инкубации включили в конце января 1996 г., постепенно увеличивая температуру воды до 5,4 °С. Личинки вылупились в феврале при 6,9–7,0 °С. До середины апреля их содержали при 12 °С, а затем температуру выровняли с естественной. Из-за аварии личинки из икры лужских и невских производителей были объединены. В конце мая мальков перевели из основного цеха в бассейны цеха в Ивановском, а к середине июня – в садки. В конце октября, когда сеголеток вернули на зимовку в основной цех, их численность составила 3950 шт., средний вес – 18,0 г.
4-я генерация (1996 г.), как и 2-я, получена из икры невского лосося. О происхождении следующих генераций маточного стада судить затруднительно, так как ЛПЭЛРЗ со времени пуска в эксплуатацию почти ежегодно ввозил икру и, иногда, молодь с других р/з (Мурза, Христофоров, 2000б). Только за время существования маточного стада количество доставленной икры лосося составило: с Невского р/з – 251,6 тыс. шт. (генерации 1995, 1996, 1997 гг.), с Нарвского р/з – 865,9 тыс. шт. (генерации 1996, 1997, 1998, 2000, 2001 гг.), из Финляндии («Лаукаа») – 100 тыс. шт. (генерация 1994 г.)* и сеголеток с Нарвского р/з – 204,5 тыс. шт. (генерации 1999, 2000, 2002, 2005, 2008 гг.). Производственные партии молоди, полученной из икры проходных производителей лосося разного происхождения, выпускали в р. Лугу и её притоки.
С 1999 г. их дополнили выпусками потомства маточного стада. Возвращавшихся в реку проходных производителей при отлове единообразно считали «дикими лужскими». Фактически, маточное стадо сформировано на основе межпопуляционной гибридизации.
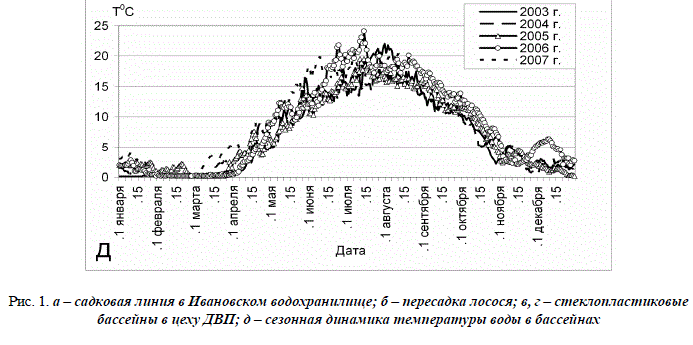
Условия содержания рыб. К началу формирования стада жилого лосося по «финской» технологии ЛПЭЛРЗ располагал только инкубационно-выростным (основным) цехом в Поречье с бассейнами 2 х 2 и 4 х 4 м. Вода подавалась из р. Хревицы насосами. Из-за сбоев в их работе, газопузырьковая болезнь нередко становилась причиной гибели молоди. На время ремонта насосов лосося переводили в садки из безузловой дели (3 x 4 x 4 и 6 x 4 x 5 м), которые устанавливали на понтонах в Ивановском водохранилище (рис. 1 а). Дважды в месяц садки промывали, так как дель обрастала нитчатыми водорослями. По мере роста молоди размер ячеи увеличивали с 3 до 10 мм. Каждый раз рыб пересаживали (рис. 1 б). Для борьбы с ихтиофтириозом, триходинозом, хилодонеллёзом, диплостомозом и миксобактериозом, наблюдавшимися у лосося в садках, проводились комплексные лечебные мероприятия. Вода в водохранилище летом прогревалась у дна до 17,0 °С, а в поверхностном слое – до 24,0 °С. В бассейнах цеха, где рыб содержали в остальное время года, водообмен был ниже нормативного и варьировал от 0,14 до 0,60 л/сек. Таким образом, первые генерации маточного стада лосося, определившие его репродуктивный потенциал на период до начала 2000-х гг., формировались в относительно неблагоприятных условиях. С весны 1996 г. часть рыб перевели в цех в Ивановском, где была организована самотёчная подача воды из водохранилища, а основной цех летом 1997 г. закрыли на реконструкцию. В ходе её намечали до 1998 г. переоборудовать также цех ДВП для содержания маточного стада лосося, от которого планировали ежегодно собирать 500 тыс. шт. икры. Фактически, реконструкция этого цеха и прокладка к нему водовода, обеспечивающего самотёчное поступление воды, завершились к концу 2000 г. Были установлены стеклопластиковые бассейны 9 х 9, 6 х 6, 4 х 4 и 2 х 2 м (рис. 1 в, г), а также система дегазации воды. Стадо переместили в цех ДВП в 2001 г. Сезонная динамика температуры воды в выростных сооружениях этого цеха представлена на рисунке 1 д. В 2006 г. смонтирована установка замкнутого типа с терморегуляцией для инкубации икры и подращивания молоди до веса 1,0 г. Она позволила получать личинок и мальков из икры лосося маточного стада в ранние сроки, но дальнейшее их выращивание при низких естественных температурах воды вызвало затруднения.
_____________
* Икра от финского маточного стада «невской линии».
Выживаемость за выдерживание и подращивание личинок была ниже нормативной. Сеголетки финского лососи лучше «приручались», но уступали потомству проходных невских производителей в устойчивости к заболеваниям и росте. При выпуске годовиками в апреле 1996 г. средний вес финского лосося составлял 14,0 г, а невского – 17,5 г. Ранее привозившаяся в 1990 г. икра от финского маточного стада имела диаметр около 4,0 мм и слабые оболочки. Большинство личинок погибло до перехода на внешнее питание, а оставшиеся особи выпущены в р. Лугу сеголетками.
Корма и добавки. Использовали гранулированные корма фирм: «EWOS» (Швеция), «Rehuraisio» (Финляндия), а позднее «BioMar» (Дания). Размеры гранул и нормы кормления выбирали по таблицам. Добавляли витамины А, С, Е, раствор CaCl2, говяжью кровь, селезёнку, салаку, а в последние годы также биостимуляторы и адаптогены – «Пуриветин» и «Мидивет».
Методы оценки биологических показателей жилых и проходных самок. Длину тела рыб измеряли по Смитту, а затем расчисляли по чешуйным препаратам их размеры к концу каждого предшествующего года жизни по стандартной формуле: Lf = Lt x Sf/St, где Lf – длина в конкретном возрасте, Lt – длина при поимке, Sf – расстояние вдоль большой оси чешуи от центра до края и St – расстояние от центра до «зимнего кольца», соответствующего конкретному возрасту (Atlantic …, 1992). Об особенностях роста судили по изменениям длины и веса тела в онтогенезе, сезонной динамике среднесуточных приростов, а также размерно-весовым характеристикам при достижении половой зрелости.
Размеры самок лосося маточного стада в конкретном возрасте сравнивали с таковыми проходных самок-производителей, отловленных для целей разведения в реках Нарве и Неве, так как популяции этих рек служили основными донорами при формировании проходной лужской популяции и стада на ЛПЭЛРЗ. Происхождение проходных самок («заводские» или «от естественного нереста») оценивали комбинированным методом, основанным на изучении внешних признаков и образцов чешуи (Antere, Ikonen, 1983). Возраст при поимке и при достижении половой зрелости определяли по чешуе и обозначали согласно правилам ИКЕС. Повторно созревающих проходных самок идентифицировали по наличию «нерестовых марок» на чешуе и исключали из выборок, принимая во внимание иные параметры роста и разрушение краевых участков чешуи. Термины «возраст», «возрастная группа» применяли, согласно Allan, Ritter, 1977, для указания общего числа прожитых рыбой лет, а также комбинации длительности пресноводного и морского периодов её жизни.
Группы проходных самок, для сравнения с жилыми самками из стада, формировали по принадлежности к тем же, либо близким генерациям.
Под «генерацией» понимали совокупность особей, происходящих из икры, заложенной на инкубацию в конкретный календарный год. Количество исследованных самок указано в таблицах. Сравнивали также репродуктивные показатели жилых и проходных самок-производителей: темп оогенеза, возраст при наступлении половой зрелости, периодичность созревания, размеры икры и плодовитость. Для этого определяли вес рыб до и после отцеживания. Измеряли и взвешивали по 50 шт. ненабухших икринок от каждой самки, вычисляли средние значения их диаметра, веса и вариабельность. Рабочую плодовитость рассчитывали по количеству икринок в стандартной навеске 50 г и общему весу отцеженной икры без овуляторной жидкости. Анализировали окраску тела, целостность чешуйного покрова и плавников. Биологические пробы включали по 6–10 самок каждой генерации. После вскрытия этих рыб оценивали состояние внутренних органов визуально и на гистологических препаратах, окрашенных гематоксилином по Гейденгайну (Ромейс, 1954). Яичники взвешивали и вычисляли гонадосоматический индекс (ГСИ). Оогенез подразделяли на периоды и фазы, а развитие яичников – на стадии, согласно ранее разработанным периодизации и шкалам зрелости (Мурза, Христофоров, 1991).
При обозначении фаз периода превителлогенеза использовали сокращения: ФЦР – фаза цитоплазматического роста, ФКВ – фаза формирования кортикальных вакуолей, ФЖВ – фаза формирования жировых вакуолей. Средние значения показателей, стандартные ошибки и достоверность различий между группами рассчитывали с использованием программы «Statistica 6».
Результаты и обсуждение
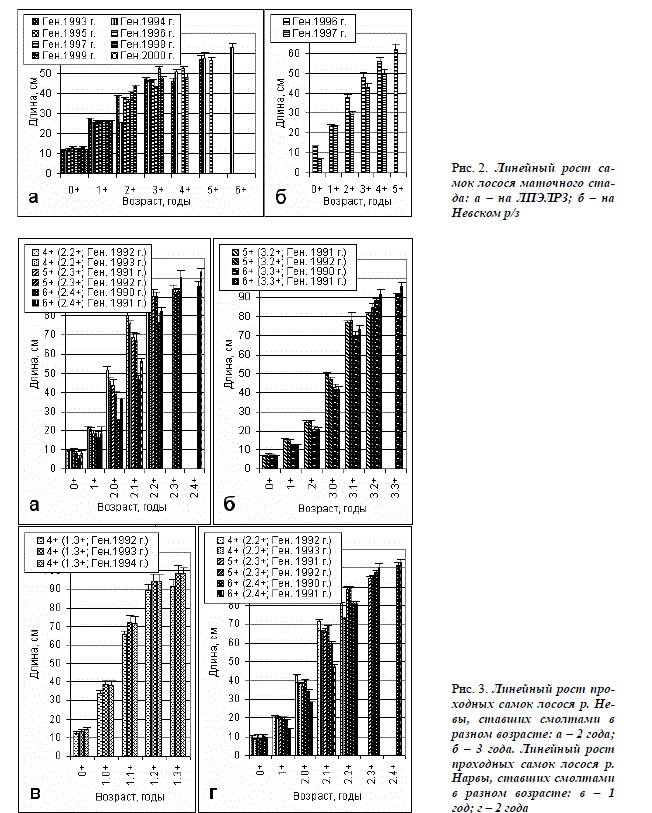
Линейный и весовой рост. Средняя длина тела жилых самок к концу 1-го года жизни (в 0+) на ЛПЭЛРЗ (рис. 2 а) и на Невском р/з (рис. 2 б) близка к таковой проходных самок
невской и нарвской популяций, имеющих заводское происхождение (рис. 3 а-г). К концу 2-го года (в 1+) самки из маточных стад иногда даже крупнее проходных, мигрировавших из рек в возрасте 2 и 3, но уступают мигрировавшим в 1 год. В возрасте 2+ жилые самки сопоставимы по длине с проходными самками обеих популяций к концу 1-го сезона нагула в море (А.0+), независимо от возраста при миграции из рек (от 1 до 3 года), а также от общего возраста, складывающегося из длительности пресноводного и морского периодов жизни (от 1.0+ до 3.0+). Динамика линейного роста лосося маточного стада на ЛПЭЛРЗ близка к таковой рыб стада на Невском р/з (рис. 2 а, б). На обоих заводах в возрасте 5+ ÷ 6+ длина тела жилых самок составляет около 60 см. Они соответствуют, либо уступают по размерам проходным самкам к концу 2-го сезона нагула в море (А.1+). Быстрорастущие жилые и проходные самки претерпевают смолтификацию и становятся половозрелыми раньше, чем тугорослые. Замедление роста у созревающих особей сопряжено с началом вителлогенного роста ооцитов. При наступлении половой зрелости длина тела жилых самок варьирует в диапазоне 31,9–45,0 см, а у проходных самок из рек Нарвы, Невы и Луги, с морским периодом от 2+ до 4+ она составляет обычно 65,0–107,0 см. Нетипично мелкие проходные самки лосося длиной 45,0–54,0 см, созревающие после 1 года жизни в море (2.1+ и 1.1+) появляются в этих популяциях после выпуска крупных смолтов (Христофоров, Мурза, 2005).
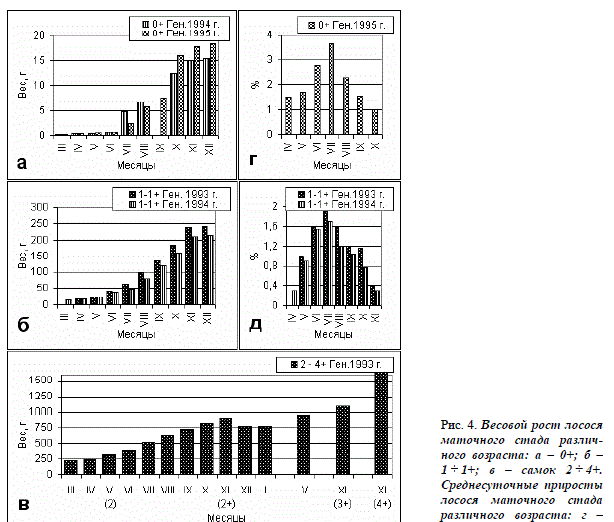
При одинаковом общем возрасте жилые производители достоверно мельче (P<0,01), чем проходные. Есть существенные различия в линейном (рис. 2 а, б; 3 а – г), а также в весовом росте (рис. 4 а, б) между рыбами разных генераций. Динамика весового роста жилых самок генерации 1993 г. в возрасте от 2 до 4+ представлена на рис. 4, в. Среднесуточные приросты у лосося маточного стада изменяются по сезонам и с возрастом. Они больше в 1-й год жизни, чем во 2-й, а максимальные значения этого показателя у рыб обеих возрастных групп отмечены в июле (рис. 4 г, д).
Оогенез и овуляция. Сроки анатомической, цитологической дифференцировки пола и динамика последующего развития половых желёз у лосося зависят от температурного режима в периоды инкубации икры, выдерживания и подращивания личинок, а также основного выращивания молоди. В яичниках мальков 1-й генерации маточного стада ооциты 1-й ступени ФЦР появились с конца апреля – начала мая. К концу мая выявлены ооциты 2-й ступени ФЦР. Их диаметр увеличивался с 85–100 мкм в начале июня до 110–168 мкм к ноябрю – декабрю. В июне – начале июля следующего года (в 1+) ооциты достигали в развитии 3-й ступени ФЦР (около 200 мкм), и их дальнейший рост в пределах этой ступени продолжался до второй зимовки. С двухгодовалого возраста самок можно было подразделить по состоянию яичников и значениям ГСИ (рис. 5) на две группы.
У особей 1-й группы весной и в начале лета темп развития старших ооцитов резко ускорялся. Они достигали 4-й ступени ФЦР (около 360 мкм), затем ФКВ, ФЖВ периода превителлогенеза и переходили к вителлогенезу (1100–1200 мкм). Продолжительность III ранней и III поздней стадий зрелости яичников у жилых самок лосося оказалась меньше, чем у проходных из Балтийского моря, созревающих в возрасте А.2+ и А.3+ (Мурза, Христофоров, 1982; Христофоров, Мурза, 1985). Значения ГСИ быстро увеличивались в октябре – ноябре (рис. 4). Усиливался половой диморфизм, хотя с высокой вероятностью (более 80%) мы отличали самок от самцов по форме головы и размерам жирового плавника ещё в возрасте 1+. Брачный наряд жилых производителей напоминал таковой проходных, но был выражен в меньшей степени (рис. 6 а, б). Тело у созревающих самок становилось вальковатым, окраска боков и жаберных крышек – тускло-серебристой, а затем свинцово-серой. Жилые производители были небольшого размера (рис. 6 б, в). Длина тела жилых самок при созревании в возрасте 2+ варьировала от 31,9 до 45,9 см (средняя 38,8±6,97 см), вес – от 400 до 1200 г (средний 737,7±132,45 г.). Яичники IV стадии зрелости занимали почти всю брюшную полость, семенники – до 60–70% (рис. 6 г). Овуляция у жилых самок происходила в середине ноября, тогда как у проходных – с середины октября. Созревание части самок лосося маточного стада на 3-м году жизни соответствует ранее разработанной нами схеме, согласно которой, при определенных условиях половая зрелость наступает на следующий год после смолтификации (Мурза, Христофоров, 1991). Для оценки влияния остаточной икры на повторное созревание, часть самок отцедили и пометили. На инкубацию заложили 25 тыс. шт. икры. Диаметр ооцитов следующей генерации в ноябре – январе составлял 750–1000 мкм (ФЖВ), редко до 1400 мкм (начало вителлогенеза). К маю следующего года (в возрасте 3 года) яичники отцеженных и неотцеженных особей были на VI-III поздней стадии зрелости. Значения ГСИ варьировали от 0,61 до 1,97%. Диаметр вителлогенных ооцитов достиг 1100–1400 мкм.
Остаточные икринки встречались в полости тела на протяжении лета и осени. Отцеженные и неотцеженные самки созрели повторно (в 3+) без пропуска цикла, в то время как у многих проходных самок переходу к повторному созреванию предшествует пропуск репродуктивного цикла (Мурза, Христофоров, 2000).
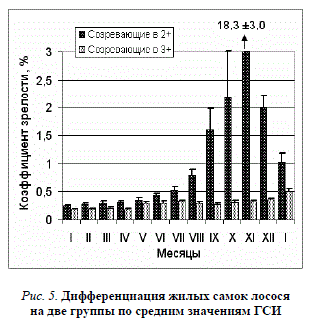
Самки 2-й группы, не созревшие осенью в возрасте 2+, имели в то время длину от 38,0 до 46,0 см, вес – от 545 до 1110 г. Они характеризовались серебристой окраской и более высоким телом. Ооциты в яичниках были мельче, чем у особей 1-й группы: до 750–800 мкм (ФЖВ) в ноябре – декабре и 1000–1100 мкм (переход к вителлогенезу) в мае следующего года. Средние значения ГСИ увеличивались в 3-й год жизни с 0,18 до 0,51% (рис. 5), а в мае 4-го года составляли 0,53±0,021%. Половая зрелость у этих самок наступила в возрасте 3+. Соотношение численности самок 1-й и 2-й групп (созревших в 2+ и 3+) в генерации 1993 г. составило 1 : 1. Среди самцов той же генерации 40% были половозрелыми в 1+ и 96,5% – в 2+. В 1997 г. от впервые и повторно созревших жилых самок генерации 1993 г. получено 108,0 тыс. шт. икры. В следующих генерациях стада, подвергшихся меньшей акселерации за счёт подогрева воды, созревание самок происходило преимущественно в возрасте 3+ (до 80–90%), реже – в 4+. Они подразделялись по состоянию гонад на две группы, как и рыбы генерации 1993 г., но продолжительность идентичных фаз и периодов развития ооцитов была больше. У большинства самок к зиме, за год до наступления половой зрелости, состояние старших ооцитов соответствовало завершению ФЖВ; вителлогенез начинался весной, а не летом, и ко времени овуляции икра оказывалась крупнее, чем у самок, созревших в возрасте 2+. Икру от самок, созревавших в 2+ (2-й и 4-й генераций – 1994 и 1996 гг.), закладывали на инкубацию также в 1997 и 1999 гг. В 2000 г. в стаде оставалось ещё 48 самок генерации 1994 г. (5+), а в 2001 г. – 9 шт. (6+). От 8 из них икру получили в 4-й раз. Даже без акселерации за счёт подогрева воды темп созревания у илых
особей выше, чем у проходных. Проходные самки из рек Луги, Нарвы и Невы лишь изредка становятся половозрелыми при общем возрасте 3+ (возрастная группа 2.1+), а обычно – в 4+, 5+ и 6+ (возрастные группы 2.2+, 3.2+, 2.3+, 3.3+, 2.4+). Кроме того, проходные производители голодают после захода в реки (Мурза, Христофоров, 2005), а жилые питаются перед сезоном размножения и после его завершения. За время зимовки они теряют брачный наряд и становятся похожими на вальчаков. «Нерестовые марки» на чешуе жилых самок выражены слабо. В результате перехода на круглогодичное содержание маточного стада в цехе р/з, серьёзной проблемой стало очень позднее и асинхронное созревание самок. Овуляция у них нередко происходит в конце ноября – декабре, когда качество спермы у самцов ухудшается и температура воды не оптимальна. Приходится стимулировать овуляцию синтетическим аналогом гонадотропин-рилизинг гормона – сурфагоном (Михелес и др., 2002). Известно, что получаемые в результате такой стимуляции яйца лосося не всегда имеют удовлетворительное рыбоводное качество (Crim et al., 1986; Taranger et al., 1992). Мы предлагали продлевать период функциональной зрелости гонад самцов лосося, используя энантат и пропионат тестостерона (Christoforov, Murza, 1996), но кардинальное решение проблемы видим в изменении условий содержания производителей обоего пола. Самки 1-й генерации стада, которых содержали в садках при естественно изменявшейся продолжительности светового дня, созревали «дружнее», чем особи следующих генераций в цехе, так как свет – один из важнейших экологических факторов, регулирующих созревание лосося (Христофоров, Мурза, 1988, 1998а). В ранних работах успешно получали икру от жилых производителей этого вида, содержавшихся в открытых прудах, бассейнах и садках (Day, 1885; Otterstrǿm, 1933; Svärdson, 1949; и др.). «Финская» технология станции «Лаукаа», положенная в основу РБО для ЛПЭЛРЗ, также предусматривает использование земляных прудов для содержания маточных стад. Станция располагает 72 такими прудами (Эскелинен, 1991а). Наряду с естественным освещением, в аквакультуре лосося широко используются также регулируемые световые режимы, позволяющие активно управлять сроками созревания производителей (Adams, Thorpe, 1989; Haux et al., 1991; Hansen et al., 1992; Björnsson et al., 1994; Oppedal et al., 1997, 2003; Taranger et al., 1998,1999; Endal et al., 2000).

Плодовитость, икра и личинки. Средняя рабочая плодовитость жилых самок лосося в возрасте 2+ на ЛПЭЛРЗ составляет 2055,8±134,4 (при индивидуальных вариациях 1000–2760) шт., в 3+ – 2573,9±107,3 шт. и в старшем возрасте – около 3 тыс. шт. Она значительно ниже (P<0,01), чем у проходных самок основных возрастных групп (табл. 1, 2).
Различия меньше в пересчёте на 1 кг веса, но около 85% жилых самок-производителей, использованных целей разведения за годы эксплуатации стада, весили меньше 2,0 кг, а проходные – в несколько раз больше. Средние значения диаметра и веса икры у жилых самок на ЛПЭЛРЗ, как и на Невском р/з, увеличиваются с возрастом (табл. 3), но остаются достоверно ниже (P<0,01), чем у проходных самок невской популяции, за исключением возрастной группы 2.1+, и у проходных самок нарвской популяции, за исключением групп 2.1+ и 2.2+ (табл. 1, 2).
Как и несколько десятилетий назад (Привольнев и др., 1964), икра у невского лосося крупнее, рабочая плодовитость меньше, чем у нарвского того же возраста. Иногда различается и цвет икры. Эти особенности могут быть обусловлены местами нагула в Балтике.
Величина коэффициента вариации веса 1 икринки у жилых самок 2+ составила 10,0–23,7 (в среднем 16,3±0,89)%, тогда как у проходных самок – 3,6–4,3% (в среднем 4,0±0,12)%. Отношение наибольшего значения диаметра икры к наименьшему соответствовало у жилых 1,6–3,6 (в среднем 2,1±0,11), а у проходных – 1,09–1,34 (в среднем 1,14±0,04). Доля нормально развивающихся эмбрионов при инкубации икры жилых самок 2+ в среднем – 76,0%, 3+ – 80,6% при нормативе 85,0%. Выживаемость икры жилых самок всех возрастных групп за инкубацию – 54,1–91,0 (в среднем 77,5)%, а икры проходных самок – 71,2–99,0 (в среднем 86,5)% при нормативе 90,0%. Длина тела личинок, вылупляющихся из икры жилых и проходных самок разного возраста, сходна, но их вес с желточным мешком существенно различается.
Средние значения этих показателей у личинок из икры жилых самок 2+ составили 19,3±0,61 мм и 41,7±0,89 мг, 3+ – 19,7±0,34 мм и 69,5±0,62 мг. Длина личинок из икры проходных лужских самок – от 18,4 до 19,9 мм, вес – от 128,5 до 166,0 мг. Средняя выживаемость личинок из икры жилых самок разного возраста за подращивание – 62,1% при вариациях от 3,4 до 82,3% и нормативе 75,0%. О низком качестве икры маточного стада лосося на ЛПЭЛРЗ сообщал и И. Ю. Попов (2001). Повышенную смертность икры и личинок, получаемых от производителей маточных стад, отмечали также на других зарубежных и российских р/з (например: Jarrams, 1997; Gaston, 1988; Крамаренко и др., 2002). Отчасти она обусловлена ограниченными ресурсами желтка в мелкой икре. Личинки из крупной икры проходных самок, лучше обеспечены им и переходят к внешнему питанию позднее, при больших размерах (Привольнев, 1960). Ко времени выхода из бугров и подъёма на плав личинки лосося из крупной икры характеризуются большей выносливостью, жизнеспособностью (Einum, Fleming, 2000; Moffett et al., 2006). Лучшие рыбоводные показатели обеспечивает икра крупных самок, длиной 75–100 см, тогда как при использовании мелких самок выживаемость икры и личинок очень низка, в большинстве случаев гибель при переходе к внешнему питанию приближается к 100% (Петренко, Халяпина, 1979). Согласно действующей «Инструкции …, 1979» (С. 35), икру со средним весом менее 120 мг следует отбраковывать.
Негативное влияние на стероидогенез у производителей маточных стад, качество их гамет, процент оплодотворения икры, выживаемость эмбрионов и личинок может оказывать также неполноценность кормов по белкам, липидам, витаминам, каротиноидам, минеральным компонентам и т.д. (Torrisen, 1983; Hardy, 1985; Leray et al., 1985; Eskelinen, 1989; Izquierdo et al., 2001; Fish Nutrition, 2002). Требования к составу кормов у созревающих рыб иные, чем у молоди. Им нужно больше каротиноидов, витаминов E и C, вовлеченных в метаболизм жирных кислот и служащих антиоксидантами. При производстве коммерческих кормов природный пигмент – астаксантин, который в естественных условиях Балтийского моря лосось получает, питаясь рыбами-планктофагами, до недавнего времени часто заменяли более дешёвым синтетическим кантаксантином (ныне запрещённым), либо использовали красители «Карофилл розовый» и «Карофилл красный», содержащие лишь до 8% этих соединений. Различия в биологических свойствах астаксантина и кантаксантина могут быть ответственны за более высокую смертность потомства культивируемых лососей, чем «диких» в эмбриональный и личиночный периоды (Craik, Harvey, 1986). По нашим наблюдениям, у самок маточного стада на ЛПЭЛРЗ в одни годы икра была желтоватой, в другие – оранжевой, что соответствует цветам астаксантина и кантаксантина. Исходным материалом для синтеза печенью предшественника желтка икры – вителлогенина служат аминокислоты, образующиеся при расщеплении пищи, мышечных белков (при голодании), а также гликоген и липиды, мобилизуемые из жировых депо. У рыб маточного стада состояние печени иногда было значительно хуже, чем у возвращающихся из Балтики производителей, и молоди, прожившей несколько месяцев в реке. Визуально отмечались светлопесочный цвет и рыхлая консистенция паренхимы, а на гистологических препаратах – некроз части гепатоцитов, много жировых пустот. Аналогичные изменения наблюдали Н. А. Ефимова и И. М. Долгачёва (2002). Нарушение вителлогенной функции печени может быть причиной повышенной вариабельности размеров икры.
С кормами рыбы получают жирные кислоты и аминокислоты (Остроумова, 2001), в том числе необходимые для формирования полноценной оболочки и желтка икры (Лизенко, Нефедова, 1982;). Производители лососевых кормов иногда заменяют рыбий жир более дешёвыми растительными липидами, а рыбную муку – соевой. Возможно поэтому, в первые годы работы с маточным стадом на ЛПЭЛРЗ мы, а в дальнейшем Ю. Н. Городилов (личное сообщение), отмечали слабые оболочки икры жилых самок. Отдельные самки имели яичники нетипично малого размера, которые на гистологических препаратах характеризовались резко выраженной асинхронностью роста ооцитов и атрезией части яйцевых фолликулов.
У одной из особей с аномальными гонадами выявлена опухоль эпифиза. Не последняя роль в нарушении развития гонад у культивируемого лосося может принадлежать содержащимся в коммерческих кормах половым стероидам (Мурза, Христофоров, 2004 в). По данным Всемирной организации здравоохранения при ООН, жиры и ткани рыб, используемые для производства кормов, содержат столько токсичных соединений, что лососей, выращенных на искусственных кормах, рекомендуется есть не чаще одного раза в месяц. Возможно, некроз плавников, недоразвитие жаберных крышек, нарушение чешуйного покрова, катаракта, ухудшение состояния печени, изменения красной и белой крови, которые мы отмечали у молоди и жилых производителей лосося на ЛПЭЛРЗ (неопубл. данные), также имели трофическую природу. Развитие этих симптомов описывается при дефиците полиненасыщенных жирных кислот, витаминов и других физиологически активных веществ в кормах (Шевченко, 2005; и др.). Нами было установлено, что в крови лосося разного возраста на ЛПЭЛРЗ концентрация гемоглобина, количество эритроцитов и доля незрелых форм этих клеток (нормобластов, базофилов и полихроматофилов) закономерно увеличивались от весны к осени, а изменения лейкоцитарной формулы происходили в разные сезоны при ухудшении состояния рыб из-за болезней и стресса, вызванного нарушениями водоподачи, перевозками и т.д. Общая тенденция состояла в снижении относительной численности лимфоцитов при увеличении долей моноцитов и полиморфноядерных клеток. Аналогичные изменения клеток белой крови известны у других видов рыб после транспортировки и введения аналога кортизона (Микряков и др., 2005).
Согласно РБО, создание маточного стада должно было предотвратить распространение заболевания «М74», но вряд ли этому способствовало кормление жилых производителей фаршем из сырой балтийской салаки (Михелес и др., 2002). Именно питание проходных лососей в Балтийском море этим объектом с низкими уровнями астаксантина и тиамина (витамина В1), но высоким содержанием тиаминазы в тканях ответственно за смертность получаемых из их икры личинок от «М74» (Мурза, Христофоров, 2004б). Финны используют для кормления маточных стад лосося не салаку, а корюшку (Эскелинен, 1991а).
Роль маточного стада в поддержании популяции лосося р. Луги. В первые годы эксплуатации стада на ЛПЭЛРЗ численность самок, использовавшихся для получения икры, увеличивалась за счёт созревания рыб очередных генераций. С 1997 г. она превысила численность проходных самок. С начала 2000-х гг., когда стадо перевели в реконструированный цех ДВП, используются не менее 150, иногда свыше 200 шт. жилых самок (рис. 7 а). Икры от них собирают значительно больше, чем от лужских проходных производителей (рис. 7 б). При этом, стали использовать меньше проходных самок и количество получаемой от них икры за время существования стада снизилось.
В 2001–2002 гг. проводился только контрольный отлов лосося в р. Луге.
Первую молодь от маточного стада получили из икры, заложенной на инкубацию в 1996 г. от производителей 1-й генерации маточного стада. Средний вес сеголеток к концу сентября 1997 г. составил 11,0 г. В 1999 г. были впервые выпущены годовики, а в 2000 г. – двухгодовики, являвшиеся потомством жилых производителей той же генерации, но из икры, заложенной в 1997 г. Эти выпуски положили начало использованию стада для выполнения основной возложенной на ЛПЭЛРЗ задачи – поддержания популяции проходного лосося в р. Луге.
Суммарная численность ежегодных выпусков годовиков, происходящих из икры жилых и проходных производителей, включая привозную, установилась на уровне, близком к 100 тыс. шт. (рис. 7, в). Регистрация в низовьях р. Луги некоторого количества заводских годовиков, в том числе мелких со слабым серебрением, не свидетельствует о готовности этих рыб к морской жизни. Анализ чешуи лужских проходных производителей лосося, проводившийся нами в рамках мониторинга, показал, что менее 10% молоди, выпускаемой в годовалом возрасте, мигрирует к местам морского нагула в тот же год, а остальные – в возрасте 2, реже 3 года. Это означает, что в основной массе выпускаемые годовики не являются смолтами. Учтённый возврат в р. Лугу от выпуска 5 последовательных генераций годовиков лосося составил 0,2–0,3%, то есть оказался близким к таковому от годовалых пестряток в других реках региона (Христофоров, Мурза, 2002, 2003). Выпуски с ЛПЭЛРЗ особей в возрасте 2 ÷ 2+ за годы существования стада были незначительными и проводились преимущественно за счёт отбраковки части ремонта, в том числе карликовых самцов (0,2–3,1 тыс. шт.). Большинство двухгодовиков метили подвесными метками, снижающими, как известно, возврат в 4–5 раз.
Спорадически выпускали также сеголеток (см. рис. 7, в). В результате выпусков количество отлавливаемых в р. Луге для целей разведения проходных производителей лосося после 2000 г. не увеличилось и ни разу не приблизилось к уровню квоты на отлов их в этой реке – 90 шт. (рис. 7, г). Не решены за 15 лет и другие поставленные в РБО задачи (см. Раздел: предпосылки …). Принимая во внимание, что основным показателем эффективности работы р/з служит состояние поддерживаемых популяций рыб, следует, по-видимому, пересмотреть биотехнологическую схему воспроизводства. Показателен положительный опыт стран, являющихся нашими соседями по Балтийскому региону. В Финляндии для восстановления почти полностью утраченных в большинстве рек популяций проходных лососевых была создана сеть государственных рыборазводных предприятий.
Главным источником рыбоводного материала стали маточные стада, созданные на основе импортированной в 1972–1978 гг. икры невского лосося. Становление широкомасштабного выращивания смолтов лосося происходило в 1978–1988 гг. В результате совершенствования биотехнологии средние размеры двухгодовалых смолтов увеличились до 70–100 г. За счёт более раннего на месяц вылупления личинок при повышенной температуре воды получили также 30-граммовых годовалых смолтов. К 1987 г. выпуски смолтов лосося невского происхождения возросли с 34 тыс. шт. до 1,07 млн. шт. (Эскелинен, 1991б; и др.). Польша при воссоздании полностью утраченных популяций лосося за счёт маточных стад также ориентировалась на выращивание смолтов, обеспечивающих наибольший возврат. Регулярные выпуски смолтов в реки начались с 1994 г., и численность их превысила 0,5 млн. шт. Менее чем за 10 лет промысловый вылов лосося в реках этой страны достиг 10 тонн, а сбор икры от проходных производителей – 2,5 млн. шт. (Бартель и др., 2008).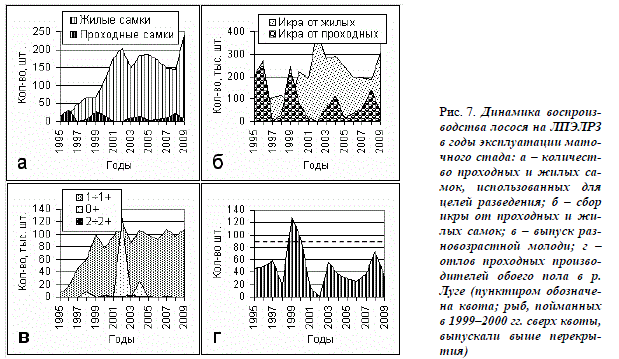
Заключение
В ходе работ, выполнявшихся на ЛПЭЛРЗ с 1995 г., показана принципиальная возможность создания в Ленинградской области маточного стада жилых производителей атлантического (балтийского) лосося и получения от него дополнительного количества гамет для целей воспроизводства. В 1996 г. впервые заложена на инкубацию икра от жилых самок.
Установлено, что темп гаметогенеза у них выше, чем у проходных особей в море, половая зрелость наступает в более раннем возрасте и при меньших размерах, а повторное созревание происходит ежегодно. Для того чтобы не усиливать скороспелость рыб, предназначенных для формирования стада, предлагается свести до минимума использование подогрева воды. Рабочая плодовитость жилых самок в несколько раз ниже, чем проходных из рек Невы и Нарвы. Они уступают проходным самкам лосося также по размерам икры и её рыбоводному качеству. Отчасти качество икры, как полагаем, можно улучшить за счёт подбора кормов. Синхронизировать овуляцию у жилых самок позволило бы содержание производителей маточного стада в летне-осенний период не в цеху, а под открытым небом – при естественной динамике освещённости и продолжительности светового дня. В целом, оценивая плюсы и минусы маточного стада лосося на ЛПЭЛРЗ, мы, как и прежде (Христофоров, Мурза, 2002) считаем нецелесообразным с биологической и экономической точек зрения долговременное содержание на региональных лососевых р/з 5–6 генераций тугорослых особей общим весом несколько тонн, требующее значительных выростных площадей, много дорогостоящих кормов и т.д. Предпочтительней использовать эти ресурсы для выращивания производственной молоди, предназначенной для выпуска в реки. Основной продукцией р/з должны быть смолты, возврат от которых на порядок выше, чем от молоди на ранних этапах развития. Для сбора икры необходимо полнее использовать доступный контингент проходных производителей лосося.
Благодарим сотрудников Отдела воспроизводства водных биоресурсов ФГУ «Севзапрыбвод» и коллектив ЛПЭЛРЗ за эффективное сотрудничество в период выполнения исследований. С большой признательностью за конструктивную поддержку вспоминаем ныне покойного директора завода Ю. И. Кузьмина.
И. Г. Мурза, О. Л. Христофоров
Санкт-Петербургский государственный университет, г. Санкт-Петербург, Россия
e-mail: Этот адрес электронной почты защищён от спам-ботов. У вас должен быть включен JavaScript для просмотра.
Смотрите также