
Биология симы и кижуча северного Сахалина
Oncorhynchus k4sutch (walbaum), обитающих в отечественных водах, изучена довольно слабо, что, видимо, связано с их относительно небольшими уловами.
Промыслово-биологическая характеристика камчатского кижуча дается в работах В.И.Гриба нова (1948), Р.С.Семі:о (1954), х.х.Зорбиди (1970); биология (1928), М.Я.Шевлевой (1951), С.Г.Соина (1954), А.И.Смирнова (1960); питанию молоди кизуча посвящены работы А.И.Сынковой (195I) и х.х.Зорбиди (1970).
Сведения о биологии симы из рец Приморья и бассейна Амура содержатся в работах А.В.Воробьева (1926), И.И.Кузне цова (1928), А.Ф.Кузнецовой (1954), П.А.Моисеева (1957).
Сима о-ва Сахалин (преимущественно южной его части) изу чалась П.А.Двининым (1952, 1957), И.С.Калмыковой (1957), А. И.Смирновым (1962), А.Н.Канидьевым (1964), С.П.Водовиком (1964); сина Камчатки и Курильских острово– Р.С.Секко (1956) и В.Н.Иванковым (1968).
в зарубежной ихтиологической литературе этим видам уделено гораздо больше внимания. Среди работ, посвященных кижучу, следует упомянуть прежде всего фундаментальное исследование Шаповалова и Тафта (Shapovalov and Taft, 1954 ), работы Хора (Ноаг, 1958 ), Фёрстера и Риккера (Poerter and R1cker, 1953 ), Хартмана (Hartman, 1965 ), Годфрея (God - frey, 1965 ) и др. Многочисленные исследования симы проводились в Японии (Tanaka, 1965; Seno and Abe, 1967; Kasaka
and Ish1da, 1969 et al. ). Кроме того, познанию симы и кижyча способствует значительный опыт искусственного разведения этих рыб, накопленный в іпонии, США и Канада.
Биология симы и кижуча Северного Сахалина практически не изучена. Некоторые сведения о размножения и развитии кижуча из р. Тамь содержатся в работе А.И.Смирнова (1960),
весовым методом содержимое 312 желудков молоди силы и 271 жeлудок кижуча. Плодовитость определена у 380 особей симы и у 95 кижучей.
Кроме собственных, в работе используются некоторые материалы Сахалинрыбвода, за любезное предоставление которых автор благодарит руководителей этой организации.
Нерестовый ход и биологические показатели производителей симы и кижуча
вопрос о пищевых взаимоотношениях молоди самы, кижуча и ке ты затрагивается в работах С.П. Воловика (1964).
Предпринимая настоящее исследование, мы ставили перед собой двоякую цель: получить представление о воспроизводстве запасов симы и кижуча в таком относительно мало исследованном районе, как Северный Сахалин, и - посмотреть, каковы ко адаптации, позволяющие сосуществовать в одних и тех же водоемах близких по образу жизни видан.
Материал собирали преимущественно в верхней и среднем течении Тыми с 1961 по 1969 г. и в верхнем и среднем течении Пороная летом и осенью 1965 и 1966 г. Количество рыб, прошедших на нерест, определяли путем непосредственного уче та на нерестилищах, а в ряде случае - при помощи рыбоучетного заграждения. Производителей для биологического анализа отлавливали неводок и вентeрeм. в годы с низкой численностью обловы вели на протяжении всего нерестового хода, в годы с высокой численностью - в течение нескольких дней во время массового хода. Молодь отлавливали различными орудиями и сачком, сеткой Киналева, мальковой волокушей и мелкоячейным не водом. Основная ее масса добыта в районе села Кировского в главным русле Тыми и крупных протоках. При обловах стремились к тому, чтобы в умовах одного дня были представлены. все mлевщиеся в водоеме возрастные группы колоди симы и ки- . хуча, к сожалению, мы не можем охарактеризовать весовой рост молоди, так как все пробы фиксировали формалином. При изучении питания содержимое желудков обрабатывали счетно-весовым методом. Чтобы дать более наглядное представление об измене ниях особенностей питания в онтогенезе, Кы располагали 'чате риал по сезонам и возрастных группах без учета года сбора, так, как если бы исследовалось одно поколение с момента по явления молоди в водоеме до ската ее в море. Для оценки степени сходства состава пищи вычисляли индекс пищевого сходства по шорьгину (1952). .
Возраст определяли по чешуе. Обратные расчисления роста производили по формуле прямой пропорциональности. Всего наки проанализировано 977 половозрелых особей (?I6 - симы и 26I - кижуча) и более 3500 шт. их молоди. Исследовано счетно
В среднем течении Тыми производители симы появляются cІ по 20 июля (чаще в I декаде июля). Массовый ход наблюдается обычно во второй половине июля и первой половине августа, заканчивается ход в период с 30 июля до 25 августа. Общая продолжительность его в различные годы - от 29 до 51 дня (табл.І).
Таблица I Сроки нерестового хода симы в среднем течении Тымы и Пороная

Примечание. Здесь и в табл.2 прочерк означает, что из-за низкой численности производителей массового хода как такового не наблюдалось.
.
Среднесуточная температура воды в реке колеблется от 6°с в июле до II°C в августе, в Поронае (в среднем течении) ход симы начинается примерно на месяц раньше, чем в Тъии, и длится на IO-20 дней дольше. Температура воды при этом колеблется от 10°с в июне до 15°с в августе.
Интервал между концом хода симы и началом хода киікуча в разные годы составляет 15-50 дней. Ход кижуча в Тъчи начинается в период с 1 по 18 сентября. Массовый ход наблюдается обычно в конце сентября - первой половине октября. Заканчивается ход между 27 ноября и II декабря, продолжаясь в разные годы от 74 до IOI дня, т.е. примерно в полтора раза дольше, чем ход симы (табл.2). Начинается ход куча при температуре 10-12°С, заканчивается примерно при іс, кассовый ход приурочен к температуре 5-2°C.
Таблица 2 Сроки нерестового хода кижуча в среднем течении

ко у самцов пределы колебаний длины были гораздо шире (+I ?I см), чем у самок (45-62 см). Средний вес симн в Тыми колебался по годам от 1950 до 2429 г, в Поронае - от 1840 до 2000 г. Самки и самцы по среднему весу не различались, но пределы колебаний веса у самцов были гораздо шире.Так,в Тыми вес самок колебался от II90 до 3550 г, вес самцов - от 750 до 3870 г.
в нерестовых стадах симы, как правило, преобладали самки (табл.4).В 1962 г. соотношение полов в Тыми было примерно І:I (9,2% самцов и 50,8% самок),в последующие годы доля самцов уменьшалась (в 1964 г. она была минимальной: 18,8%). По скольку у симы наряду с проходными самцами существуют карли ковые неогенические самцы, можно предположить, что значение их для воспроизводства вида возрастает в годы, когда уменьшается * количество проходных самцов (табл.4).
По мнению некоторых авторов (Барач, 1952; Крохин, 1967), у проходных лососей особей, развивающихся по карликовому типу, тем больше, чем выше обеспеченность пищей молоди во время пресноводного периода. Этим ,по-видимому, можно объяснить изменение доли проходных самцов из нашем случае.Дело в том, что обеспеченность пищей молоди симы в Тыми во многом зависит от численности осенней кеты - наиболее массового лосося в этой реке .Икра, теряемая кетой при нересте, является важных компонентом питания молоди симы. Трупы производителей ,в массе оставшиеся в водоемах в течение осени, зимы и весны, слухат пищей личинок ручейников, веснянок, поденок,хирoномид и других насекомых, в огромном количестве обитающих на них или в не посредственной от них близости. Личинки и взрослые формы этих насекомых составляют основу рациона молоди симы. Кроме того, лососи ,перерывая грунт при устройстве гнезд, делают более доступными для молоди симы (и других видов рыб) представите дей донной фауны.
Поколение, симы 1961 г., составившее основу численности нерестовой популяции в 1964 г., нагуливалось в первый год жиз ни при благоприятных условиях.Хорошие условия нагула были обеспечены высокой численность кеты и большими потерями ее икры во время нереста в результате аномально сильного осенне-- го паводка. Кроме того, много производителей кеты погибло в от шнуровавшихся водоемах,где трупы их сохранялись вплоть до весны следующего года.
Если массовый ход симы совпадает по времени с летней меженью, то кижуч в кассе идет на нерест в период осенних паводков, которые в отдельные годы принимают катастрофический характер из-за дождей, вызваной тайфунами.. ,
Средняя длина производителей симы в Тыми колебалась за пять лет наблюдений 07 52,5 до 55,6 см (табл.3). Примерно такой же величины была сима в Доронае (52,4-53,2 см). Средние размеры самок и самцов существенно не различались, одна
Таблица 3
Длина и вес симы и кижуча, идущих на нерест в Тым. и Поронай

Примечание: В дробях: числитель - пределы колебаний, знаменатель - средняя.
Таблица 4
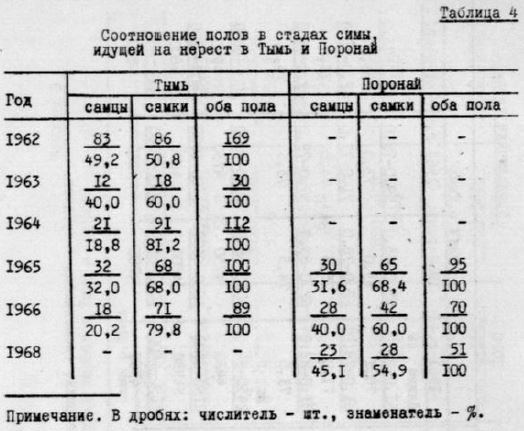
Таблица 5 Возрастной состав нерестовой популяции симы в Тыми и Поронае

Примечание. В дробях: числитель - шт., знаменатель - %.
Поколение симы 1962г. нагуливалось при худших условиях. Заход кеты был вдвое меньше, чем в предшествующем году, потери икры при нересте были небольшими, так как осенний паводок был крайне слабым. Возможно, этим объясняется увеличение до 32% доли проходных самцов в 1965 г., когда особи 1962 года рождения в массе вернулись на нерест.
Идущая в Тымъ на нерест сима представлена двумя возрастными группами: 2 + 1 32t. Последняя группа резко преобладает, составляя в разные года от 77,7 до 93,2% общей численности. Среди самцов доля четырехлетков несколько выше, чем среди самок (табл.5).
в Поронае возрастной состав симы сложнее. Там она представлена тремя возрастными группами: 211,31и 32t. Однако основу численности, как и в Тым, составляют рыбы в возрасте 3 + (от ?І,0 до 82,4%). Численность четырехлетков проживших один год в реке, составляет всего 4,3-7,8%.
Примечание. В дробях: числитель - шт., знаменатель - %.
Карликовые самцы, которых мы исследовали в одном из . притоков верхнего течения. Тыми (n = 29), были представлены тремя возрастными группами: It (48,0%), 2+ (48,0%) и 3+(4,0). Размеры двухлетков колебались от 9,2 10 10,7 см (в среднем 10,1 с), трехлетков - от 10,3 до 15,0 см (в среднем 13,3см).
Возрастные группы производителей симы из Тыми и Пороная по длине и весу, как правило, не е различаются (табл.7 и 8). Исключение составляют лишь самцы из Тыми, у которых в 1964 г. трехлетки были мельче четырехлетков, и самки из Пороная, у - которых в 1968 г. трехлетки существенно превосходили по раз мерам четырехлетков. В то же время длина при скате в море у этих возрастных групп имела большие различия , и в дальнейшем, в морской период жизни происходило выравнивание размеров за счет различного темпа роста.
Таблица ? Длина и вес различных возрастных групп симы из Тыми
В отличие от симы соотношение fодов у кижу ча" не изменялось по годам и было примерно 1:I при незначительном преобладании самок (табл.6). Нерестовая популяция кижуча в Рыми во все годы наблюдений была представлена особями одного возраста - 32t.
Таблица 6
Соотношение полов в стаде кижуча, идущего на нерест в Тымы


Таблица 8 Длина и вес различных возрастных групп симы из Пороная
Примечание. В дробях: числитель - шт., знаменатель - %.
Сравнение наших данных по возрастному составу идущих на нерест симы и кижуча с данными других исследователей показывает, что на Северной Сахалине эти виды созревают в более позднем возрасте, чем в других районах. На Хоккайдо и Южном Сахалине преобладающей возрастной группой нерестующей симы являются трехлетки (2+) (Крыхтин, 1962; Танака, 1965). кижуч Британской Колумбии и Ванкувера более чем на 95% представлен особями в возрасте 24t. Эта хо возрастная группа часто преобладает в подходах кижуча из рек Камчатки (Грибанов, 1948; Семко, 1954; Зорбиди, 1970).
Таблица 9
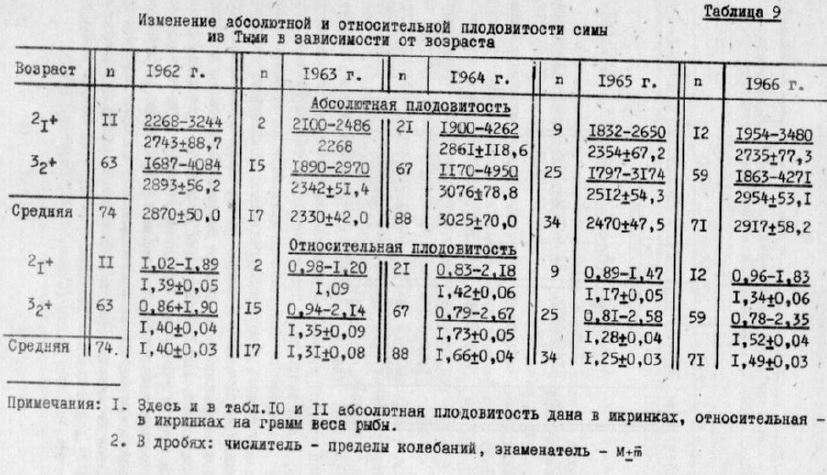
Индивидуальная абсолютная плодовитость симы в Туни колебалась от II70 до 4950 икринок, а-средняя абсолютная плодовитость - от 2300 до 3025 икринок (табл.9). Плодовитость, поронайской симы была примерно такой хе (табл.10). По этому признаку сима Северного Сахалина существенно превосходит си ку из рек южной части острова, у которой средняя абсолютная плодовитость не превышает 1770 икринок (Двинин, 1952; Крыхтин, I962). Как в Тыми, так и в Поронае плодовитость трехлетков и четырехлетков в пределах одного года различалась незначительно, в то время как различия ее по годам были достаточно ведники. При этом плодовитость в четные годы была гораздо выше, чем в нечетные. Средняя абсолютная плодовитость по четях годах колебалась от 2870 до 3025, по нечетным – от 2330 до 2470 икринок. Плодовитость рыб одного поколения, созревающих в разном возрасте, также подчинялась этой закономерности. В Тыми у симы, родившейся в четные годы, плодовитость четырехлетков бывает ниже плодовитости трехлетков. и наоборот, у поколения нечетных лет плодовитость четырехлетков выше. Аналогично изменяется и относительная плодовитость симы (см. табл.9, 10).
По-видимому, конечная величина плодовитости симы зависит от изменяющихся условий ее нагуда в море. То, что плодовитость симы выше в четные годы, когда низка численность и горбуши, служит косвенным подтверждением мнения И.Б.Бирма на (1967) о влиянии численности поколений горбуши на условия нагула, а следовательно, роста и созревания других видов лососей в морской период жизни, -
Индивидуальная абсолютная плодовитость кижуча колебалась от 1760 до 900 икринок, средняя - от 4460 до 5370. В противоположность симе абсолютная плодовитость кижуҷа в не- . четные годы выше, чем в четные. Однако у поколений кижуча сметных лет эти различия выражены слабее, чем у симы (табл.I). Относительная плодовитость кижуча в отличие от симы не обнаруживает тенденции изменяться по четным не четным годам.
Таблица 10 Изменение абсолютной и относительной плодовитости симы из Пороная в зависимости от возраста
Изменение абсолютной и относительной плодовитости симы из Тыми в зависимости от возраста

Примечания: I. Здесь и в табл. 10 и II абсолютная плодовитость дана в икринках, относительная
в икринках на грамм веса рыбы. 2. В дробях: числитель - пределы колебаний, знаменатель - М+jin
Таблица II Абсолютная и относительная плодовитость кижуча из Тыми

Нерест и развитие эмбрионов Нерест симы в Тыми происходит с конца июля до начала сентября. В Поронае нерест более ранний: с первых чисел ир дя до средину августа. Нерестилища расположены в верхнем течении основного русла и притоков. Типичными нерестовыми водоемами симы являются небольшие ручьи шириной от 2 до 5 и, с частым чередованием плесов, перекатов и ям (глубины над перекатами - 15-35 см, над плесами - 40-60 см, над ямами - до 120 см). В некоторые годы сиыа нерестилась также на участках перекатов основного русла шириной 25-40 м. Эти участки были расположены ниже верхней границы нерестилищ горбуши. Сима в подобных стадиях держалась группами, занимая нерестовые площадки в тесном соседстве с горбушей, но никогда не смешиваясь с ней. В основном русле реки в нересте принимали участие только проходные самцы, в то время как в ручьях - около одной самки держалось по несколько (в среднем 5, максимум I2) карликовых самцов. Несмотря на то, что в некоторые годы проходных самцов бывает немного, самок, нерестующих только с карликовыми самцами, нам довелось наблюдать лишь дважды. Скорость течения на нерестилищах симы в местах устройства бугров колебалась от 0,2 до 0,8 м/сек. Бугры располагались на глубине 10-25 см (табл.12). Верхняя границa зa кладки икры в буграх колебалась от 8 до 20 см, нижняя граница - от 20 до 35 см. Бугры имеют овальную форму, чаще вытянутую по течению.
Мы пытались выяснить, свойственны ли подобные колебания плодовитости симе и кижучу из других районов ареала,однако ограниченность материалов не позволила внести ясность в этот вопрос. М.Л.Крыхин (1962) сообщает, что плодовитость симы Юго-Западного Сахалина в 1953 и 1954 г. буда практически одинаковой. По данным И.И.Кузнецова (1928), плодовитость амурской симы в нечетком 1927 г. была выше, чем в кижуча из рек Большой и Камчатки колебаний по четными и нечетныи годам не обнаружила (Семко, 1954; Зорбиди, 1970).
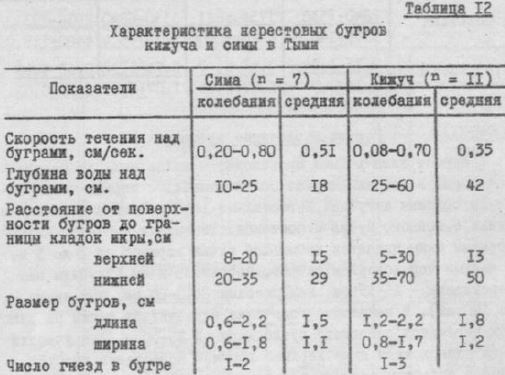
Таблица 13 Количество икры силы в нерестовых буграх ключа Кирпичного (бассейн Тыми) в сентябре 1966 г.
 в сентябре 1966 г.")
Поскольку нерест симы приходится на период летней межени, нерестовые гнезда рыба устраивает обычно в средней части русла, как правило, на прямоточных участках. Температура воды во время нереста колеблется от 8 до II°С.
Таблица 2 Характеристика нерестовых бугров куча и силы в Тыми
Скорость течения на буграми, см/сек. |o,20-0,80] 0,5 Глубина воды над буграми, см.
I0-25 I8 Расстояние от поверхности бугров до границы кладок икры, симы
верхней
В конце сентября 1966 г. (примерно через 25 дней после окончания массового нереста) мы вскрыли 7 бугров симы в ключе Кирпичном, впадающем в Тымы в верхнем ее течении.
в буграх находилось от 746 до 1227 (в среднем 983 икринок, что составляло 34% от средней абсолютной плодовитости симы в том году. В четырех буграх икра была отложена в два гнезда, в трех буграх имелось по одному гнезду. Выживаемость в момент вскрытия составляла в среднем 98% (табл.I3).
Сахалинскому кижучу свойствен наиболее ранний нерест по сравнению ск их у рек Камчатки и побережья Америки. Массовый нерест начинается в Тыми обычно во второй половине октября и длится до конца второй декады ноября. Во второй половине декабря нерест практически превращается. В январе нересту цих кижучей нам не доводилось видеть.
В сентябре и начале октября производители кижуча поднимаются в самые верхние притоки Тыми (реки Белую, Громова, Овень). Рыбы более поздних подходов распределяются по много численным протокам на участке Тыми от пос.Красная Тымь до пос.Зональное, где предпочитают устраивать гнезда на выходах грунтовых вод, используя нерестилища, на которых до этого нерестилась осенняя кета. Нерест кижуча протекает преимущественно после спада осеннего паводка. Среднесуточная температура воды от начала к концу нерестового периода понижается с 9 до 19с. Скорость течения на нерестилищах колеблется от 0,08 до 0,70 м/сек. Бугры устраиваются на глубинах от 25 до 60 (в среднем 42) см. Длина и ширина бугров кижуча по II измерения составляли соотвественно от 1,2 до 2,2 (в среднем І,8) ии от 0,8 до 1,7 (в среднем I,2) и, т.е., несмотря на более крупные по сравнению с симой размеры тела, кижуч делает бугры, мало отличающиеся по величине от бугров симы (см.
табл.I2). Икра в буграх встречалась на глубинах от 5 до 70 см. в бугре бывает от одного до трех гнезд, чаще их два.
В 1962 и 1963 г. с февраля по апрель мы вскрыли ІІ бугров кижуча. Число эмбрионов, обнаруженных в буграх, колеба лось от 5 до 1441 (в среднем 642), что составляло примерно
13% от средне абсолютной плодовитости. Выживаемость икры составляла в среднем 50%, выливаемость личинок - I00%. Общая выживаемость эмбрионов в буграх равнялась 73% (табл.14). При вскрытии бугров эмбрионы в икринках были накануне выклева: жаберные лепестки наполовину прикрывались жаберными крышками, челюсти были сформированы. Выклюнувшиеся эмбрионы имели нерассосавшиеся или рассосавшиеся на I/4 желточные мешки.
- Рост и соотношение полов у молоди симы и кижуча в речной период жизни
Таблица 14
Количество и выживаемость эмбрионов кизуча в нерестовых буграх Тыми, вскрытых в конце
февраля - начале апреля

Личинки симы начинают выходить из грунта, видимо, во второй половине апреля. Выход из грунта молоди кижуча происходит с первой половины мая до середины июня. Мальки симы, . пойманные в последних числах мая, имели длину от 32 до 38 (в среднем 33) мм и вес от 270 до 50 (в среднем 330) мг. желточные мешки у них к этому времени полностью рассосались, у, ких учей, пойманных тогда же, 57% особей имели остатки желточного мешка, вес которого в процентах от веса тела составлял от 1,3 до 20,9% (в среднем 9,4%). Длина молоди кижуча колебалась от 27 до 38 (в среднем ЗІ) мм, вес - от 150 до 550 (в среднем 260) кг. В течение июня и ирля мальки кижуча отставали в росте от симы, но уже во второй половине августа молодь обоих видов имела одинаковые размеры и в дальнейшем, вплоть до времени ската из реки, практически не различалась(рис.1). Сеголетки и двухлетки наиболее интенсивно растут с июня по сентябрь, двухгодовики - с апреля по май. с сентября до на чала декабря рост рыб резко замедляется, с декабря до начала апреля приросты их незначительны и в основном, видимо, приходятся на март.
Все сказанное относится к поколениям симы и кижуча 1961 - и 1962 г. в другие годы молодь этих видов может существенно
различаться по темпу роста. Так, в 1965 г. покатники кижуча , были крупнее покатников симы. Данные по темпу роста этих рыб, полученные путем обратных расчислений, также указывают на более крупные размеры кижуча по сравнению с симой при скате в море (табл.15 и 16). На основании этих хе материалов можно установить, что в море кижуч растет гораздо быстрей симы (см. табл.15 и 16). К концу первого года пребывания в море Киуч по длине превосходит симу на 4-6 см. Разрыв этот в дальнейшем увеличивается за счет более продолжительного морского периода жизни у кижуча.
При вскрытии бугров кижуча мы дважды находили в них икру кеты, отложенную ранее. Кроме того, дважды при вскрытии бугров кеты обнаруживали, что они переконань кижучем. Поскольку глубина закладки икры у кижуча больше, чем у кеты, видимо, большая часть икры кеты при перекопке погибает. Возможность конкурентных отношений из-за нерестилищ между кижучем и другими лососями предполагает и И.И.Кузнецов (1928). Однако в настоящее время при низкой численности кижуча на Сахалине подобная конкуренция невелика.

Таблица 16
Рост кижуча (в ск) в Тыми (по данным обратных расчислений)

Таблица 15
Примечание. В дробях: числитель - длина, знаменатель -
прирост. Соотношение полов у молоди обоих видов иное, чем у производителей в нерестовых подходах. Води у симы, идущей из ко ря, значительно преобладают самки,то у молоди, правда, очень не значительно - самцы. Некоторое преобладание самцов наблюдает cя" также у пестряток кижуча (табл.І?) .Среди покатников кижуча в июле 1965 г. соотношение полов было примерно II (48,5% самок и 5,5% самцов; n=I07). У покатников симы преобладали самки (63% самок, 37, самцов; n=86).
самки
Рост симы (в см) из Тыми и Пороная (по данным обратных расчислений)
Тымы, 1963 г.
Таблица 17 Соотношение полов и возрастов (в %) у молоди симы и кижуча в различных участках верхнего течения Тыми
 у молоди симы и кижуча в различных участках верхнего течения Тыми")
Примечание. В дробях: числитель - длина, знаменатель - прирост.
Определить возрастной состав симы и кижуча в той или иной реке в целом трудно из-за широкого распространения молоди по многочисленных ключам и протокам, неодинаковой плотности их заселения и возрастных различий в предпочитаемых местах обитания. Однако сравнение возрастного состава молоди в разных стадиях и дает дать представление о расселении ее по реке. Расселение молоди сокы заканчивается в основном в течение первого дета. Часть сеголетков весной и в начале дета скатывается из нерестовых ручьев в основное русло и главные протоки. Так, во второй половине сентября 1969 г. количество сеголетков в нерестовом ключе Кирпичном и в про токе Крутиловской, где сима не нерестует, было одинаковым (табл.18).
 и кижуча (0) в верхнем течении Тыми (наблюденные данные)")
Рис.І. Линейный рост молоди симы (а) и кижуча (0) в верхнем течении Тыми (наблюденные данные): I - поколение 1961 г.; 2 Епоколение 1962 г.
Сеголетки кижуча в первый год расселяются преимущественно по старицам и протокам основного русла. В ручьи они заходят в первое лето в меньшем количестве, о чем свидетельствуют данные о возрастном составе молоди кижуча в разных стадиях (табл.18).
Серебрянки обоих видов в верховьях реки начинают встречаться в большом количестве с конца мая, однако скат происходит гораздо позже. В среднем течении Тыми покатников симы и кижуча нам довелось наблюдать в 1963 и 1965 г. лишь со второй декады июля. В пробах покатников, отловленных в июле 1965 г., симa нa 24,4% была представлена двухлетками (І+) и на 75,6% трехлетками (2+) (n=86/. У кижуча двухлетки со (ставляли 10,3%, трехлетки - "89,7% /2 =107/.
в 1965 г. покатники кижуча несколько превышали по длине покатников симы. Двухлетки кижуча имели длину II,9 си, тех летки - 13,8 см, у симы двухлетки были длиной II,2 см, трех
Примечание. В дробях: числитель - самцы, знаменатель - самки.
Скат симы и кижуча на Северном Сахалине происходит в более старшем возрасте, чем во многих других районах ареала, где основная масса молоди покидает реки на Втором году жизни. Это обусловливает сравнительно поздний возраст их полового созревания на севере острова.
Если в летние месяцы возрастной состав молоди симы и кижуча;
групп и появления сеголетков, численность которых в разные го Ды бывают различной , то в период зимовки (с ноября по апрель) возрастной состав должен быть относительно постоянным.Зимой 1962 и весной 1963г. дважды были обновлены большие скопления молоди симы и кижуча в крупной протоке - Первая Тьмъ. За четыре месяца возрастной состав симы не претерпел каких-либо заметных изменений, у кижуча 39 количество годовиков уменьшилось на II,3% (cir.табл.18).Поскольку хищников в этой протоке не было, элиникацию годовиков кижуча мы склонны объяснять гель кинтозами, которым кижуч, по нашим наблюдениям, подвержен гораздо больше, чем сима.
Питание симы и кижуча и их пищевые отношения с другими видами рыб
Таблица 18
Возрастной состав (в %) молоди симы и кижуча в различных участках верхнего течения Тыми
 молоди симы и кижуча в различных участках верхнего течения Тыми")
Питание молоди симы и кинуча в течение пресноводного периода сходно и по сезонной динамике, и по качественному составу корма. Наибольшие индексы наполнения хелудков отме чаются в летние месяцы и в сентябре (рис.2). с октября до
апрель они снижаются примерно в 2-3 раза. Наименьшая напод - -ненность желудков наблюдается у этих рыб в период ската -
в июле. С июня по сентябрь в пище- всех возрастных групп обо “ их видов доля воздушных и наземных насекомых составляет 40- 15
50% по весу. Среди воздушных насекомых преобладают '1mago двукрылых, поденок и муравьев, среди донных организмов - личинки и куколки хирoномид, поденок, веснянок, ручейников, водяных хуков и бабочек. По мере- роста рыб в их питании заметно уменьшается доля личинок хирoномид и возрастает доля личинок ручейников. Значение личинок веснянок и поденок остается относительно постоянным на всем протяжении пресно водного периода. Важным пищевых компонентом молоди обоих видов является икра тихоокеанских лососей, которую они на чинают потреблять уже на первом году хизни. В желудках сеголетков симы икра встречалась с средины августа, у сеголетков кижуча - сначала октября. Корм этот в течение зимних и весенних месяцев доминирует у обоих видов, однако для кижуча его значение несколько выше. Потребление икры - продолжается вплоть до конца июня следующего года. -
- Покатная молодь кеты и горбуши единично встречается в желудках симы и кижуча в течение июня и июля. Среднее количество экземпляров в желудке по разным пробам не превышает 0,2.
Значительно сходство между симой и кижучем также во "внутрисуточной ритмике пищевой активности их молоди в прод
покатной .-период. Оба вида имеют два подъема пищевой активно сти - с 7 до II часов и с 13 до 21 часа (Гриценко, Ардави чус, 1967).
Протока Первая Тымъ Вид | Пол |2.XII.1962 г. | 4.II.1963 г.
оба
Все сказанное опитаник исследуемых видов относится к
верхнего течения. Для изучения особенностей питания симы и 7 кижуча в разных стациях во второй половине сентября 1969 г.
были собраны пробы в ручье (ключ Кирпичный), ключе (протока, Крутиловская, и старице (Малый Кувет), расположенных в окрестностях села Кировсќаго. Та хе был собран материал для изучения питания ряда массовых видов рыб, обитбюрдих совместно с симой и кижучем: гольца (Salvel1nus alpinus (L.), девятииглой колошки (Pung1tluѕ рung1t1.us tymens1s (Make, сибирского гольца (Nemachilus Barbatulus toni (Dyъ.). Параллельно была определена доля различных видов рыб вих, тиомассе исследуемых водоемов. Для удобства сравнения про бы обдих видов гольцов и колюшки подразделялись на размер- ! но-возрастные группы, соответствующие размерам различных возрастных групп симы-и кижуча. Так, Родец в возрасте 1+1 примерно равен по -длино, сеголеткам симы и кижуча; в возрасте 1 и 3+ - двухлеткам, а особи более старшего возрас та - трехлетнам. Сибирский голец подразделен на две размерные группы: -менее 10 см и более 10 см, что соответствует длине
симы и кижуча в возрасте 0+ и It. Колюшка в нашей пробе со - ответствует по-чдлине сеголеткам лососей.
"В ключе Кирично более 50% веса пищевого комка молоди симы и кижуча составляли представители авиафауны (за исключением сеголетков симы, у которой они составляли лишь 42%). В питании двухлетков дихуча воздушные беспозвоночнно имели наибольшее значение (68,3%). Среди них преобладали по во су комары, поденки, муравьи и муҳи. Среди донных беспозвоночных превалировали личинки жуков и бабочек. Личинки веснянок, поденок и ручейников, вместе взятые, обычно не превышали по весу I0%. Таким образом, включе Кирпичном питание сеголетков симы и кижуча имеет больше сходства, чем питание двухлетков, которые значителѣно различаются по доле аваифауны в пищевом комке (рис.3, табл.19).
")
Рис.2. Сезонные особенности питания молоди симы (а) и кижуча (0) в верхнем течении Тыми: І - икра лососей,2 - молодь лососей,3 - личинки хирономид,4 - личинки поденок,5 - личинки веснянок,6-ли чинки ручейников,7 - личинки жуков,8 - личинки бабо чек,9 - водяные хуки,10 - прочие водные организмы, ІІ - комары,I2 - поденки,13- муравьи,I4 - мухи,15 - прочие воздушные и наземные насекомые
сибирскерии хижу саме голец" Калю шля
Голец по характеру питания отличается от силы и кижуча больше, чем последние друг от друга. В первую очередь это от носится к молоди наименьших размеров, которой более, чем лососям, свойственно бентическое питание. Из донных организмов Голец гораздо больше, чем сима и кижуча, потребляет личинок ручейников и веснянок. в желудках симы и кижуча из протоки Крутиловской оказалось вдвое меньше воздушных насекомых, чем в желудках рыб этих видов из ручья. Личинки жуков и бабочек по-прежнему играли ваную роль в питании симы и кижуча, но вместе с тем резко возросло значение личинок ручейников, веснянок и поденок. в питании гольца преобладали донные организмы (96%), в основном личинки ручейников (59%) и веснянок (18%). Различия в питании симы и кижуча в ключе оказались существен нее, чем в ручье, но и в ключе такого рода различия между этими видами были все же меньше, чем между каждым из них и гольцой (табл. 20).
Таблица 20 Индексы пищевого сходства симы и гольца и кижуча


Рис.3. Процентный состав пищи молоди симы, кижуча, гольца, сибирского гольца и девятииглой колюшки в различных стациях верхнего течения Тыми во второй половине ,
сентября 1969 г.: І - личинки хирoномид; 2- личинки ручейников; 3 - личинки поденок; 4 - личинки веснянок; 5 - личинки жуков; б- личинки бабочек; 7 - жуки; 8 - ракообразные; 9 - рыба; 10 - прочие водные организмы; ІІ - кошары ; 12 - поденки ; 13 - муравьи; 14 - мухи; 15 - прочие воздушные насекомые
Таблица 19 Индексы пищевого сходства симы, кижуча и гольца из ключа Кирпичного

Примечание. При вычислении индексов пищевого сходства в
табл.19, 20, 2I мы не диференцировали воздушных и наземных насекомых по групах.
В старице, как и в ручье и ключе, значительную часть пищевого комка , лососей составляли воздушные насекомые (преимущественно мухи). Из водных организмов у всех возрастных групп обоих видов лососей преобладали жуки, второе место занимали личинки жуков, третье - личинки хирономид..
Вместе с молодью лососей в старице обитают сибирский голец и девятииглая колкутка. Главным кормом сибирского голь ца являлись личинки хирoномид (до 73% по весу): В* питании колошки основными компонентами были ракообразные - циклоны и харлактициды (47%), а также личинки хирон омид (42%). Ли чинки хуков и бабочек, служащие кормом симы и кижуча, в ze
лудках сибирского гольца и колюшки встречались в небольшом - количестве, а воздушные насекомые вообще не попадались.
Таким образом, и в старице сима и кижуч по характеру питания отличаются друг от друга меньше, чем от соседствующих видов - сибирского гольца и девятииглой колодки (табл.2I). Подобное расхождение спектров питания исключает напряженность пищевых отношений между молодью лососей и сорянки, рыбами летом и в начале осени, когда имеются воздушные насекомые, однако с октября по май такая напряхенность весьма вероятна.
Таблица 21 Индексы пищевого сходства симы, кижуча, девятииглой колошки и сибирского гольца
в старице Малый -Кувет

 и воздушных и наземных (2) организмов (в % от общего числа съеденных) в желудках молоди рыб")
Рис.4. Соотношение водных (I) и воздушных и наземных (2) организмов (в % от общего числа съеденных) в желудках молоди рыб, пойманных в различных стациях верхнего течения Тыми во второй половине сентября 1969 г.
Весовой состав пищевого комка достаточно хорошо показывает, какой корм является основным в питании того или иного вида, однако для анализа пищевых отношений небезинте добывании различных груп кормовых организмов. Для этого мы вычислили долю воздушных и водных организмов в процентах от общего числа организмов, обнаруженных в желудках (токе было проделано в отношении других исследованных видов рыб во всех трех стациях). Оказалось, что в ключе Кирпичном молодь симы и кижуча активнее охотится за воздушны
и насекомыми, чем за водна и организмами. При этом сего дети и двухлетки кижуча проявляют большую активность, чем эти же возрастные группы симы. Но особенно активны в этом отношении карликовые самцы симы (2+) /рис.4/.
Активность гольца в добывании авайфауны увеличивается по мере роста, но в целом она все 20 гораздо больше направлена на добывание бентических организмов, что отличает его от симы и кижуча. В протоке Крутиловской по сравнению с ключом Кирпичным сима и "кууч активнее охотятся за бентическими организмами, чем за воздушными и наземними; двухлетки обоих видов потребляют воздушных насекомых больше, чем сего летки. Соотношение количества этих групп организмов в желудках симы и кижуча весьма сходно. Голец по сравнению с ними выступает как типичный бентофаг, в пище которого воздушные насекомые составляют лишь 5% от общего числа съеденных организмов. В старице у син и кижуча воздушные насекомые со ставляют немного менее половины съеденных организмов, а обитающие там совместно с ними сибирский голец и колюшка, как уже было сказано, совершенно не потребляют воздушных насекомых
Р).
Таким образом ,различия в характере питания симы и кижуча из разнотипных водоемов несущественны, тогда как те же различия между каждый из этих видов и соседствующими сними гольцами и девятииглой колюшкой довольно значительны.
Сходство в питании симы и кижуча наводит на мысль о возможности пищевой конкуренции между ними. Косвенным подтверждение тому может служить уменьшение численности обоих видов в местах перекрывания их ареалов. Но существуют адаптации, позволяющие им ослаблять напряженность отношений. Основной адаптацией является значительное расхождение мест обитания в пределах того или иного водоема. Так, В.И.Гриба нов (1948) отмечал, что молодь кижуча "избегает мест с быстрым течением и держится главным образом по тихим и глубоким плесам ключей, речных стариц и проток и под нависшимя берегами основного русла реки". Молодь симы гораздо реофильнее, она избегает стариц и заросших заводей. Кроме того, как это установил С.П.Воловик (1964), мальки симы и кижуча в разное время используют прибрежные мелководья рек вследствие разных сроков выхода из грунта. Существенное расхождение в местах обитания молоди кижуча и стальноголового лосося, который в экологическом отношении может служить аналогом симы, обнаружил Хартман (Hurtman, 1965) в реках Северной Америки.
Наши материалы также подтверждают значительное расход дение предпочитаемых мест обитания молоди симн и кижуча. Так, в ключе Кирпичном биомасса молоди кижуча возрастает 0 верховьев книзовьям, составляя от II до 28% всей био массы рыб. Биомасса молоди симы, наоборот убывает от верховьев книзовьям, составляя от 56 до 38% (рис.5). В целом в этом водоеме биомасса симы в два с лишних раза превышает биомассу кижуча. В протоке Крутиловской сима составляет 57, аких уч 32% ихтиомассы. Молодь обоих видов образует в этом водоеме единое скопление. В старице молодь симы была пред ставлена единично (2% общей биомассы рыб), а кажуч, наобо рот, являлся массовым видом.
 в, ихтиокассе")
Рис.5. Доля различных видов рыб (в %) в, ихтиокассе
некоторых водоемов верхнего течевия Тыми: аб,в - соответственно верхнее, среднее и аижнее течение ключа Кирпичного, - про тока Крутиловская, д - старица Малый Сувет; І-- Киуч, 2 - cима, 3 - Голец, 4 - сибир ский годец, 5 - девятииглая колюшка, 6 сахалинский таймень, 7 - гольян
Численность симы и кижуча на Северном Сахалине
Сима и них уч на Дальнем Востоке считаются второстепенными объектами лососевого промысла на том основании, что их численность значительно уступает численности горбуши якеты. Однако сопоставление численности симы и кижуча с численностью европейских лососей, промыслу и, разведению которых уделяется большое внимание, опровергает это традиционно сложившееся мнение. Так, если уловы семги на Кольском полу острове в 60-е годы колебались от I,6 до 3,9 тыс. в год, то вылов одного только кижуча на Камчатке в эти же годы превышал 30 тыс .д. и, хотя сима и кижуч менее ценны, чек атлантический лосось, все же следует всячески способствовать увеличению их численности.
неурожайных лет не прослеживается. Может быть такого чередо вания и не существует, а может быть его нарушает японский морской промысел.
В некоторые годы в ряде районов значение симы и кижyча в промысле бывает довольно велико. Так, в 1950 г. на западном побережье Сахалина в смешенных уловах с горбушей сима составляла 35-40% (Двинин, 1952). В р.Тунин (Приморье), по данным Главрыбвода, в довоенные годы выдавливалось 7 8 тыс. симы; А.В.Воробьев (1926) отмечает, что наибольшие ее уловы в этой реке приходились на годы с низкой численностью горбуши.
в реках северо-восточного побережья Сахалина кизуч многочисленнее симы. За годы наших исследований наиболее массовый заход его в Тни отмечен в 1961 г., когда на нерестилищах верхнего течения было учтено около 40 тыс .произ водителей. В последующие пять лет численность его была гораздо ниже и колебалась от 6 до 15 тыс.шт. Минимальный за ход - меньше I тыс. шт.-был в 1967 г. (табл.22).
Таблица 22 Количество производителей симы и кижуча, учтенных на нерестилищах верхнего течения Тыми (в тыс.т.)
")
на Дальнем Востоке - должно предусматривать искусственное разведение симы и кижуча. Правда, разведение этих видов сопряхе но с некоторыми трудностями, связанных с особенностями по лучения зрелых производителей и содерканием колоди в течение длительного пресноводного периода. Но сима и кижуч имеют ряд преимуществ перед традиционными объектами рыбоводства на Дальнем Востоке - көтой и горбушей. Так, сима и кижуч, выпущенные в двух-, трехлетнем возрасте, обеспечат более высокий и стабильный возврат, чем горбуша и кета. Обостренный инстинкт родной реки и сравнительно позднее наступление дошадости позволяет вести промысел симы и кижуча в устьях рек, разумно регулируя пропуск и изъятие.
• Барач Г.П. Значение ручьевой форели в воспроизводстре запасов черноморского лосося (кухи). - "Зоол.Хури.", т.3I, . вып.6, 1952, с.906-915. Бирман И.Б. О межвидовых отношениях тихоокеанских лососей в море. - Изв.ТИНРО, 1.57, 1967, с.3-24.
• Воловик С.П. Материалы по биологии колоди симн Oncorhyn
chus masu (Brevoort) некоторых рек Сахалина. - "Вопр.
ихтиол.", т.3, вып.3, 1953, с.506-512. Воловик с.п. Пищевые отношения молоди симы с молодыю дру
гих лососевых - в реках Сахалина. - Изв.ТИНРО, т.55,
1964, с.83-96. Воробьев А.В. Промаҫед симы и его развитие. - “Болл.рыбн.
нет св.
Наибольшая численность симы наблюдалась в 1962 г. (на год позze максимального захода киҗуча), когда на нерести лица прошло 14-I8 тыс. производителей. В последующие годы она колебалась от 10 до 1-2 тыс.шт. Данных за 1961 и 1967 г. нет, но, по некоторым сведениям, сима тогда была крайне ма лочисленной. Интересно отметить, что наибольшие заходы симы наблюдались в четные годы - 1962 и 1964, когда численность горбуши была низка, что согласуется с упомянутыми выше наб людениями Воробьева. Однако четкого чередования урожайных и
Грибанов В.И. Куч. – Изв.ТИНРО, 1.28, 1948, с.45-02. Гриценко О.Ф., Ардавичус А.И. осуточной ритмике пищевой
активности некоторх хидных рыб в связи с выеданием молоди дальневосточных лососей. - Co.НТИ (ВНИРО), вып.7, 1967, с.27-3I.
36
Двинин П.А. Лососи Южного Сахалина, - Изв.ТИНРО, т.37;
1952, с.70-I08. Двинин П.А. Места зимовок симы в районе Сахалина. - ПРыбн.
хоз-во", 1957, E II, с.79-8I. Зороиди х.х. Питание. молоди куча в некоторых водоемах
Камчатки. - Изв.ТИНРО, 1.73, 1970, с.72-87. Зорбиди х.х. Одинамике стада к уча.“- Изв.fИНРО, 1.78,
1970, с.6I-72. Иванков В.Н. Тихоокеанские лососи о-ва Итуруп (Куриль
ские острова). - Изв.ТИНРО, т.65, 1968, с.49-74. Невлева М.Я. Морфология и темп развития тихоокеанских до
- сосем. - Изв.ТИНРО, 1.34, 195I, с.I23-130. * Калмыкова М.С. Питание и некоторые особенности биологии
симы. - "Рыбн.х-во", 1957, E.8, с.59-60. Канидьев А.Н. Условия нереста и развитие кры симы Oncor
1964, с.289-292. Крохин Е.Н. Материалы к познанир карликовой красной Oncor -
hynchus. nerka (walb.) в Дальнем озере (Камчатка).
"Вопр. ихтиол.", т.7, вып.3, 1967, с.433-445. Круҳтин .Л. Маeтриалы о речном периоде хизни симн. -
Изв. ТИНРО, 3.48, 1962, с.84-32. Кузнецов И.И. Некоторые наблюдения над размножением амур
ских и камчатских лососев. - Изв.Тихоок-научн.-пром.ст.,
т.2, вип.3, 1928, 196 с. Кузнецова А.Ф. Некоторые данные по биологии колоди симн. -
Изв.ТИНРО, 1.4I, 1954, с.337-340. Моисеев П.А. Образованке хилой формы симы (Oncorhynchus
masu morpha formosanus (Jordan et Oshima) Boaccene
р,Седанки. - ДАН СССР, т.II2, 1957, EI, с.163-164, Смирнов А.И. К характеристике биологии размножения и раз
вития кижуча - Oncorhynchus kisutch (albaum).
- Вестник МГУ, cep.УІ, биод., вып.I, 1960, с.9-20. Смирнов А.И. Экология размножения симы Oncorhynchus mази
(Brevoort) - ДАН СССР, т.143, 1962, E6, с.29-3I. Сеико Р.С. Запасы западнокамчатски лососей их промысло
Бое значение. - Изв.ТИНРО, 1.4I, 1954, с.3-I09,
Семко Р.С. Новые данные о западнокамчатской симе. - "Зоол.
. курн.", 1.35, вып.7, 1956, с.IOI7-022. Соин С.Г. Закономерности развития детней кеты, горбуши и
сицы. - Tp.coвeщ. по вопр. осос,хозва Дальн.Вост.м., изд.
АН СССР, 1954, с.144-155: Сынкова А.И. О питании тихоокеанских лососей в камчатских
водах. - Изв.ТИНРО, т.34, 1951, с.I05-122. Elорыгин А.А. Питание и пищевые взаимоотношения рыб Кас
пийского моря. М., Пищепромиздат, 1952, 268 с. Chapman, D. 1962. Aggressive behaviour in Juvenile,coho
salmon as a cause of emigration. J.Fish. Res.Ba.Canada.
Vol.19, No.6, pp.1047-1080. Foerster, R. and Ricker,w. 1953. The coho salmon of Cultus
lake and Sweltzer Creek. J.Fish. Res. Bd.Canada. Vol.10,
No. 6, pp.293-319. Godfrey, H, 1965. Salmon of the North Pacific Ocean. Pt.9,
Cpho salmon 'In offshore waters. Bull. Int.N.Pacif.'F1sh.
Comm. No. 16, pp.1-40. Hoar,w. 1958. The evolution of migratory behavior among
juvenile salmon of the genus Oncorhynchus. J.Fish. Res. -ва .Canada. Vol.15, No !3, pp.391-428. . Hurtman,G.F. 1965. The role of behaviour in the ecology and in
teraction of underyearling coho salpon (Oncorhynchus
kisutch) and steelhead trout (Salmo gairdneri). J.Fish.
Res.ва.Canada. Yон.22, No.4, pp. 1013-1081. . Kosaka,s. and Ishida, T. 1969. A note on the ecology of
juvenile masu salmon, Oncorhync Xus' masu in a brook. * Bull.Pac.P1sh.Hokka1do. Vol.20, No.2, pp.65-74. 1.1ster,D.в. аnd Walker,С.В. 1966. The effect of rlos control
on freshwater survival of chum, coho, and chinook salmon
in the B1g Qualicur River. Can.F18h.Cult. No.37, PP.3-26. Lister, D.B. and Genoc,H.S. 1970. Stream habităt utilization
by cohabiting underyearling of chinook (Oncorhynchus . - 'tschawytscha) and cobo (0.kisutch) salmon in the B18
Qualicum River, British Columbia. J.F1sh.Res.Bd.Canada,
Vol.27, No.7, pp.1215-1224. Mund1e,J.H. 1969. Ecological implications of the diet of
Juvenile coho’in streams. Symposium of Salmon and Trout in Streams. H.R.MacMillan Lectures in Fisheries. Univ.Brit.Columbia, pr.135-152.
e4sher 1єв:
39
Pritchard, A. 1940. Studies on the age of the coho salmon
(Oncorhynchus kisutch) and the spring salmon (Oncorhynchus tschawytscha) in British Columbia. Trans.
Roy.Soc.Can., Ser.3, Vol.34, Sect.5, pp. 99-120. Sano,s. and Abe,S. 1967. Ecological study of the masu salmon
(Oncorhynchus masu (Brevoort).. The observation on the Smolt in the coastal waters. Sci.Rep.Hokkaido Salm.
Hatch. No.21, pp.1-10. Shapovalov,L. and Taft,A. 1954. The life histories of the
steelhead rainbow trout (Salmo gairdneri gairdnert) and silver salmon (Oncorhynchus kisutch). With special reference to Waddell Creek, California and recommenda tion regarding their management. Fish.Bull., Sacra
mento, No.98, 375 pp. Tanaka,s. 1965. Salmon of the North Pacific Ocean. Pt.9,
A review of the biological information on masu salmon (Oncorhynchus masu). Bull. Intern.N.Pacif.Fish.Comm. No.16, pp.75-135.
Редактор К.Д.Косова Отдел научно-технической информации
д- 96170
Подписано к печати - зо/ш - 73 г. Формат 92,8х50
Заказ No 42 Тираж - 100экз. Объем 2,0 п.л.
Цена 20 коп. Фоторотопринтний цех ВНИРО Москва, Б-140, Верх.Красносельская, 17
Триенко В.ф.