
ПИТАНИЕ МОЛОДИ КЕТЫ (ONCORHYNCHUS KETA) В ПЕРИОД ЛЕТНЕГО НАГУЛА В ЮЖНОЙ ЧАСТИ ОХОТСКОГО МОРЯ
Траловая съемка по учету молоди лососей была проведена в Охотском море в районе шельфа и свала глубин юго-восточного Сахалина, в прол. Лаперу-за, зал. Анива, на акватории открытой части моря над абиссальной котловиной и над шельфом и свалом глубин у о. Итуруп и северной оконечности о. Кунашир
(см. рисунок). Для облова молоди лососей использовали разноглубинный канатный трал 54,4/192 м. Длительность каждого траления составляла 30 мин. В прол. Лаперуза выполнено 9 станций, в зал. Анива — 17, на шельфе и свале глубин у юго-восточного Сахалина — 31, в открытых водах южной части Охотского моря — 11, у южных Курильских островов — 37 станций. Всего за период рейса выполнено 100 траловых станций, 21 из них результативная, на которых отловлено 158 экз. молоди кеты. Для анализа питания были взяты все отловленные экземпляры кеты, из них в районе Сахалина — 25 шт., в районе южных Курильских островов — 133 шт.
Схема тралений (точки), величина улова за траление (разноразмерные кружки) и изотермы на поверхности моря (прерывистые линии) во время проведения учетных работ по молоди лососевых на НПС "Дм. Песков" в конце июня — июле 2003 г.
Scheme of trawl stations, catch per trawling of fry salmon and surface isotherms counting works on the research vessel "Dmitry Peskov" during end of June — August, 2003
Обработку материала проводили по методике, предусматривающей индивидуальную обработку каждого желудка. На начальном этапе определяли наполнение желудка по растянутости стенок и количеству пищи (Методическое пособие ..., 1974) и степень переваренности каждой группы или вида (Чучукало, На-пазаков, 1999). Для выяснения значимости отдельных видов и групп видов в пищевых трактах молоди кеты вычисляли следующие количественные показатели: общий индекс наполнения желудков (%оо), процентный состав пищи по массе, частоту встречаемости кормовых объектов. Для выяснения значимости кормовых компонентов в питании использовали коэффициент обилия (КО) (Палий, 1961): КО = В х ЧВ, где В — средняя относительная биомасса, %; ЧВ — частота встречаемости данного вида. Индексы сходства спектров питания вычисляли по формуле, предложенной Шенером (Schoener, 1970): Cxy = 100 — — 0,5Е( \ px — Py\), где Cxy — индекс сходства питания видов x и y, %; px, py — доля конкретного компонента соответственно в рационе видов x и у, %.
Суточный рацион рассчитывали с использованием метода, учитывающего время переваривания различных кормовых групп (Чучукало, Напазаков, 1999). Для сравнения с теоретическими показателями рассчитали величину суточного рациона по балансовому равенству, учитывающему обменные процессы (Вин-берг, 1956): Р = П + Т + Н, где П и Т — количество пищи, используемое на прирост и обмен; Н — неусвоенная часть пищи.
При использовании данного равенства нами были сделаны некоторые допущения, применены унифицированные коэффициенты и температурные поправки (Сущеня, 1971): усвоение пищи — около 80 % пищи (зоофаги), средняя скорость весового суточного прироста — 4,45 % (Справочные материалы ..., 2000). Величину энергетического обмена в зависимости от массы рыбы определяли с учетом температурной поправки для приведения значений обмена к средней температуре в конце июня — июле (9-13 оС). Для перехода к суточным показателям трат на обмен использовали оксикалорийный коэффициент, равный 4,86 кал/мл О В полученную величину также внесли поправку на калорийность пищи и самой рыбы (Кизеветтер, 1954; Остроумова, 2001).
Гидрологическая обстановка
Пространственная структура полей температуры и солености характеризовалась наличием квазистационарного антициклонического вихря в южной части зал. Анива, серией вихрей циклонической и антициклонической направленности над шельфом и свалом глубин у юго-восточного Сахалина. На большей части зал. Анива и в прол. Лаперуза температура поверхности моря (ТПМ) составляла 11,0-12,5 "С и только в северо-западной части зал. Анива, в районе апвеллинга, она была ниже — 3,0-4,5 " С (см. рисунок). На шельфе у юго-восточного Сахалина и в открытых водах над абиссальной котловиной ТПМ изменялась от 9,3 до 13,0 "С. Изотерма 3 "С, считающаяся нижней температурной границей для обитания молоди лососей, в прол. Лаперуза и в северной части зал. Анива располагалась на глубине 20-25 м, выходя на поверхность в районе ап-веллинга и заглубляясь до 35 м в центре антициклонического вихря. На шельфе и свале глубин у юго-восточного Сахалина изотерма 3 "С находилась на глубине 20 м, поднимаясь до 15 м в зоне циклонических вихрей.
На севере о. Итуруп в зал. Простор ТПМ изменялась от 4-5 "С в его южной части до 8-9 "С на периферии залива, изотерма 3 "С залегала на глубине 20-30 м. Южнее, в районе заливов Курильского и Куйбышевского, ТПМ была выше — 7-10 "С, изотерма 3 "С находилась на глубине 15-20 м. Южная оконечность о. Итуруп была под влиянием адвекции тихоокеанских вод через прол. Екатерины, ТПМ составляла 4-5 "С. Изотерма 3 "С находилась на горизонте около 20 м. Над шельфом северной оконечности о. Кунашир ТПМ была выше — 8,1-11,3 "С, изотерма 3 "С залегала на глубине 25-40 м.
По данным стандартных океанологических разрезов ситуация отличалась от типичной сверх нормы положительными аномалиями солености в слое 050 м на разрезе мыс Свободный — море, что, видимо, связано с более развитым прибрежным апвеллингом и положительными аномалиями температуры воды в верхнем 50-метровом слое на разрезе мыс Терпения — море, что может указывать на ослабление прибрежного апвеллинга.
Распространение и биологические показатели молоди кеты
Встречаемость молоди кеты в прол. Лаперуза, зал. Анива и над шельфом у юго-восточного Сахалина составила соответственно 22,2, 5,9 и 18,2 % от числа выполненных станций. Попадалась она штучно — всего в этих районах выловили 11 экз. кеты. В открытых водах Охотского моря ее облавливали исключительно в западной части абиссальной котловины вплоть до траверза мыса Терпения на севере. Встречаемость в данном районе составила 45,5 % при общем вылове 14 экз. Средний улов на усилие по данным результативных тралений возрастал от внутренней части шельфа (1,3 экз./трал.) к открытым водам Охотского моря (2,8 экз./трал.).
В районе южных Курильских островов встречаемость молоди кеты составила 24,3 %. Основные ее скопления были обнаружены в зал. Простор (33 экз.) и на шельфе северной оконечности о. Кунашир — в обширном районе,
прилегающем к прол. Екатерины (91 экз.). В зал. Простор она была распространена вдоль шельфа до северной оконечности острова, но основной улов пришелся на станции, находившиеся на траверзе р. Рейдовой. Молодь кеты штучно облавливали и в открытых водах над глубинами более 200 м на внешней границе зал. Простор (6 экз.) и на траверзе прол. Екатерины (3 экз.).
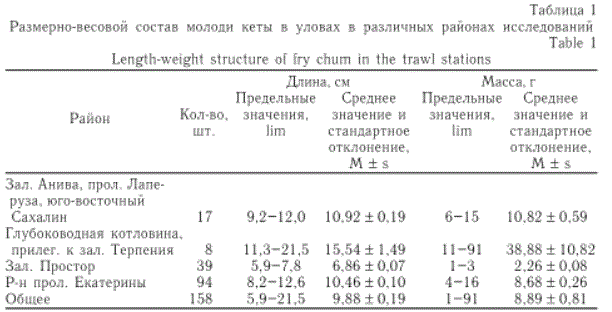
По размерно-весовым показателям выделялась молодь кеты, выловленная в координатах 47°21' с.ш. 144°59' в.д. На этой станции в улове находилось три экземпляра кеты длиной от 15,0 до 20,3 см и массой тела около 55 г (табл. 1). Длина и масса тела молоди в юго-западной части Охотского моря и на шельфе северной оконечности о. Кунашир были сходны и изменялись соответственно от 8,2 до 12,6 см и от 4 до 16 г. Наиболее мелкая кета была обловлена в зал. Простор (о. Итуруп). Длина рыб изменялась в пределах 5,9-7,8 см, масса тела — 1-3 г.
Таблица 1
Размерно-весовой состав молоди кеты в уловах в различных районах исследований
Кета, желудки которой подверглись анализу, была выловлена над глубинами, превышающими 40-45 м. В результате обработки желудков молоди кеты, обитающей в южной части Охотского моря, идентифицировано 34 формы морских беспозвоночных, икры, личинок рыб и воздушных насекомых из 10 крупных таксономических групп (табл. 2). Наиболее разнообразно представлена группа ракообразных, состоящая из 12 видов копепод, 2 видов гипериид, 7 форм эвфаузи-евых и 3 личиночных форм десятиногих раков. Рыбы представлены личинками из 3 семейств — стихеевых, скорпеновых, керчаков. В большом количестве в желудках обнаружены ойкоплевры. По основному показателю "востребованности" пищи — коэффициенту обилия — мы можем охарактеризовать видовой состав пищевых объектов в питании молоди кеты для каждого выделенного географического района.
В прол. Лаперуза и у мыса Анива (см. рисунок) в питании молоди кеты встречено 14 видов и форм пищевых объектов различной систематической и экологической принадлежности: наземные насекомые; прибрежные, неритичес-кие, эпипелагические, эврибатные планктонные беспозвоночные ракообразные и рыбы. Из общего количества выделено пять предпочитаемых пищевых компонентов с коэффициентом обилия > 100: эвфаузииды (Euphausia pacifica, Thy-sanoessa longipes), гиперииды (Themisto japónica), личинки рыб и копеподы (Neocalanus plumchrus). По относительной доле в пищевом комке (ПК) преобладали гиперииды, по частоте встречаемости (практически в 50 % случаев) — личинки рыб (табл. 2).
Таблица 2
Основные кормовые объекты (относительная биомасса, частота встречаемости и коэффициент обилия) в питании молоди кеты летом 2003 г.
в южной части Охотского моря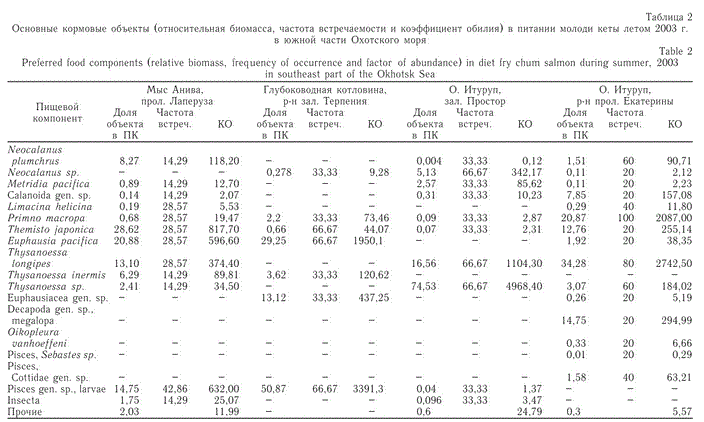
Питание молоди кеты над глубоководной частью Охотского моря, прилегающей к зал. Терпения, менее разнообразно. Общее количество кормовых организмов меньше, чем в прибрежном районе (7 видов), причем это в основном интерзональные виды, характерные для открытых вод Охотского моря. По коэффициенту обилия в желудках преобладали молодь рыб и эвфаузиевые раки (Euphausia pacifica, Thysanoessa inermis, Euphausiacea gen. sp.), остальные группы и виды соответствуют второстепенной пище.
У южных Курильских островов — в зал. Простор (о. Итуруп) и в районе прол. Екатерины — в желудках кеты обнаружено всего 29 форм кормовых объектов. В зал. Простор по коэффициенту обилия преобладали эвфаузиевые раки рода Thysanoessa (скорее всего Th. longipes) и копеподы рода Neocalanus. В районе прол. Екатерины в питании доминировали четыре группы кормовых объектов (КО > 100): эвфаузииды (Thysanoessa longipes), гиперииды (Primno macropa и Themisto japonica), личинки десятиногих раков на стадии мегалопа и копеподы (Calanoida gen. sp.), принадлежащие, скорее всего, к роду Neocalanus. К второстепенной пище во всех исследованных районах относятся крылоногие моллюски, щетинкочелюстные и ойкоплевры.
В отличие от ранее проведенных исследований, гиперииды Themisto japonica не были преобладающей пищей, частота их встречаемости максимально достигала 67 %, а в прикурильских водах не превышала 33 % (Сынкова, 1951; Андриевская, 1975). Также следует отметить, что основную часть этого вида составили экземпляры длиной от 1,5 мм и выше.
При сравнении спектров питания молоди из разных районов наибольшее сходство (на уровне 40 %) отмечено для юго-восточного Сахалина (прол. Лаперу-за, мыс Анива) и глубоководной части моря, прилегающей к зал. Терпения, и на уровне 33 % — между водами южного Сахалина (прол. Лаперуза, мыс Анива) и районом прол. Екатерины. Наименьшее сходство кормовых объектов установлено для глубоководных вод в районе зал. Терпения и зал. Простор о. Итуруп — 0,5 %.
В желудках были отмечены организмы длиной от 0,65 до 45,0 мм. Самые мелкие формы представлены оболочниками, крылоногими моллюсками, мелкими калянидами; крупные формы — половозрелые эвфаузииды и молодь рыб. Кета размером 5 см уже начинает потреблять довольно крупные объекты длиной до 15 мм (при средней длине 6,25 мм). С увеличением размеров кеты спектр значительно расширялся: в питании сохранилась высокая доля мелкоразмерных жертв (Themisto japonica, Oikopleura vanhoeffeni), но возрос верхний предел размеров кормовых объектов (Chaetognatha, Thysanoessa).
Единственная наиболее заметная смена пищевых групп (по размерам) отмечена при длине кеты около 12 см. Практически полностью из питания исчезают мелкие оболочники, каляниды, молодь гипериид. Средняя длина потребляемых организмов возрастает до 22-45 мм за счет крупных видов эвфаузиид (Th. inermis, Th. longipes, Euphausia pacifica), а также молоди рыб.
В литературе неоднократно отмечалось, что мелкие формы в питании кеты встречаются на протяжении практически всего морского периода жизни, а при массовом развитии в планктоне абсолютно преобладают в питании — до 99100 % массы ПК. В прибрежных водах это относится в первую очередь к мелким копеподам (в том числе Harpacticoidae), которые у разноразмерных групп молоди могут составлять до 97 % массы пищи (Каев и др., 1993). В морских открытых водах такой группой являются гиперииды (Сынкова, 1951; Андриевская, 1975). Такие же закономерности отмечены и для крупной мигрирующей кеты (Сынкова, 1951).
Таким образом, с ростом увеличиваются средние и максимальные размеры жертв, а также разнообразие пищи, но при обилии "излюбленной" пищи молодь предпочтет скорее ее, чем более крупную, но малочисленную добычу. Если учесть, что в Охотском море (район центральной глубоководной котловины и при-
брежные воды Сахалина) в июле происходит основной нерест Themisto japónica (Чучукало и др., 1999), то наличие и доминирование в питание этой группы вполне объяснимо и закономерно.
В целом, молодь кеты питалась довольно интенсивно, все рыбы имели наполнение желудков более 1 балла. Средний индекс наполнения желудков (ИН) составил 178,15 %оо, изменяясь по участкам от 38,84 до 333,06 %оо. Наименьший ИН отмечен в глубоководной части моря, прилегающей к зал. Терпения, наибольший — в зал. Простор (о. Итуруп) (табл. 3).
Таблица 3
Основные количественные показатели питания молоди кеты летом 2003 г.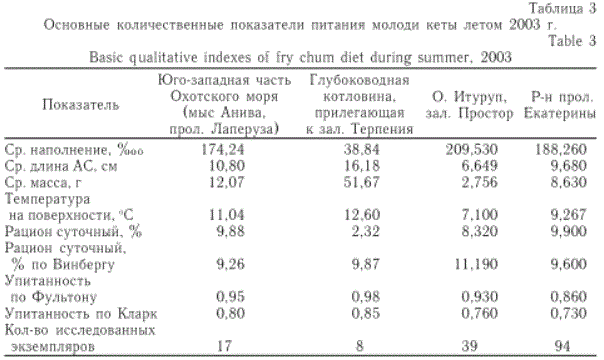
Степень переваренное™ пищи в желудках была выше 50 %. В желудках с высоким ИН одновременно присутствовала и свежепроглоченная, и полупереваренная, и полностью переваренная пища. И лишь в утренние часы (4-5 ч) практически вся пища была свежей (на 85-90 %). В это время наполнение желудков было минимальным, и лишь около 8-12 ч наблюдается некоторое увеличение ИН. Второе менее заметное увеличение количества свежей пищи отмечено в период с 15 до 17 ч, и в это время происходит хорошо выраженное возрастание накормленности рыб.
По степени переваренности пищи в течение всего периода наблюдений нами была определена скорость переваривания основных кормовых объектов молоди кеты. Для гипериид и личинок декапод среднее время переваривания составило ~ 4 ч (в пределах от 1 до 8 ч). Время переваривания эвфаузиид — в среднем 6,2 ч (от 1,5 до 17,0 ч), личинок и молоди рыб — 13,4 ч (от 7,1 до 28,1 ч), копепод — 6,1 ч (от 2 до 17 ч).
Факт такой высокой скорости переваривания весьма интересен. По данным К.М.Горбатенко, В.И.Чучукало (1989), средняя скорость переваривания ракообразных у лососей составляет от 8 до 12 ч. В летний период это время значительно сокращается по нескольким причинам: гиперииды представлены мелкими экземплярами, сам процесс переваривания происходит намного эффективнее у молоди, чем у взрослых рыб, более высокая температура воды также ускоряет обменные процессы в организме (Винберг, 1956).
Таким образом, для ракообразных время переваривания в среднем составило 6,15 ч, или 0,26 сут, для рыб — 13,4 ч, или 0,56 сут.
Рацион, рассчитанный с учетом времени переваривания пищи, варьировал в широких пределах как между отдельными экземплярами, так и по районам. В
среднем по всей акватории исследований суточный рацион составил 7,61 % массы тела рыбы, изменяясь от 2,32 до 9,90 % (табл. 3). Наиболее низкий рацион оказался у кеты юго-западной части Охотского моря (в водах, прилегающих к зал. Терпения), где отмечена самая крупная кета.
Рацион, полученный непосредственно по прямому учету потребленной пищи и времени переваривания, дает нам картину фактического питания или "потребленной" пищи.
Суточная потребность, рассчитанная с учетом темпа роста и трат на обмен (по балансовому равенству), так называемая "потребная" пища (Методическое пособие ..., 1974), составила в среднем 9,95 % массы тела рыбы. Варьирование этого показателя по районам было совершенно незначительно и отмечено в пределах от 9,26 до 11,19 % (табл. 3).
Разность значений рационов, полученных путем решения балансового равенства и прямым учетом съеденной пищи, дает величину, которая показывает, насколько в данном районе удовлетворяются пищевые потребности молоди кеты. В нашем случае мы наблюдаем значительный "дефицит" питания над глубоководной акваторией, прилегающей к зал. Терпения: он составил 76,5 % от необходимого суточного рациона. Другие районы в этом отношении можно считать вполне благополучными.
Выводы
В пищевом спектре молоди кеты идентифицировано 34 разнообразных формы морских беспозвоночных, личинок рыб и воздушных насекомых. Из них по коэффициенту обилия доминировали эвфаузиевые раки, гиперииды, личинки рыб и копеподы.
Молодь кеты потребляла организмы длиной от 0,65 до 45,0 мм. С увеличением размера кеты спектр значительно расширяется, при этом сохраняется высокая доля мелкоразмерных жертв (Themisto japónica, Oikopleura vanhoeffeni) и увеличиваются максимальные размеры жертв. Единственная наиболее заметная смена пищевых групп по размерам отмечена при длине кеты около 12 см.
Летом 2003 г. молодь кеты питалась довольно интенсивно, все рыбы имели наполнение желудков более 1 балла. Средний индекс наполнения желудков составил 178,15 %оо.
Для гипериид и личинок декапод среднее время переваривания составило ~ 4 ч (в пределах от 1 до 8 ч). Время переваривания эвфаузиид в среднем составило 6,2 ч (от 1,5 до 17,0 ч), личинок и молоди рыб — 13,4 ч (от 7,1 до 28,1 ч), копепод — 6,1 ч (от 2 до 17 ч).
Рассчитанная суточная потребность в пище молоди кеты с учетом темпа роста и трат на обмен (по балансовому равенству) составила в среднем 9,9 % массы тела рыбы. Рацион, рассчитанный методом непосредственного учета потребленной пищи, был несколько ниже и составил 7,6 % массы тела рыбы.
И.А.Немчинова, С.А.Коваленко (СахНИРО, г. Южно-Сахалинск)
Смотрите также