
Влияние условий созревания самок Азово-Черномерских кефалей: лобана, сингиля и пиленгаса на рыбоводные показатели икры
Для работ по искусственному воспроизводству азово-черноморских кефалей: лобана, сингиля и пиленгаса на НИБ ЮгНИРО «Заветное» производителей отлавливают в Керченском проливе в период их нерестовых миграций из Азовского моря в Черное. Гидрологический и гидрохимический режимы пролива крайне неустойчивы, что отражается как на динамике, так и продолжительности хода рыб.
С повышением температуры воды в Азовском море (весной и летом) ход пиленгаса и лобана в Керченском проливе интенсифицируется, в уловах возрастает число рыб с завершенной IV стадией зрелости. У сингиля, нерестящегося осенью, наоборот, интенсивность нерестового хода растет на фоне постепенного охлаждения Азовского моря с 24 до 19 °C и обычно достигает максимума в конце августа-начале сентября. В этот период результаты индуцированного созревания кефалей очень сильно зависят от направления течений, так как в бассейны, где содержатся созревающие рыбы, вода подается из Керченского пролива. Во время азовского течения в летний период условия содержания рыб на НИБ ЮгНИРО «Заветное» часто становятся близкими к таковым на местах нагула, температура воды достигает 25-26 °C. Во время же сгонных явлений в Черном море при мощном черноморском течении температура может снижаться до 9-13 °C.
В данной работе мы обобщаем многолетние экспериментальные данные (1985-2007 гг.) по исследованию влияния абиотических условий при индуцированном созревании производителей кефалей на состояние их половых желез и качество зрелой икры.
Исследование этого вопроса является одной из важных задач при искусственном воспроизводстве ценных видов рыб, поскольку совершенствование технологии преднерестового содержания производителей в контролируемых условиях гарантирует стабильное получение качественных икры и личинок.
Работы выполнялись на НИБ ЮгНИРО «Заветное», где использовали «диких» рыб, отлавливаемых в Керченском проливе, а также на экспериментальной базе ХТМО (Будакская коса, Одесская область, 1989-1990 гг.) на производителях пиленгаса маточных стад. Для экспериментов отбирались рыбы с ооцитами, достигшими IV стадии зрелости.
Производителей содержали в бетонированных или пластиковых бассейнах объемом 2,5-3 м3 при плотности 3-5 экз./м3 в замкнутых системах и в условиях постоянной проточности при аэрации воды сжатым воздухом.
Были проведены серии экспериментов, в которых производителей кефалей инъецировали гомогенатом ацетонированных гипофизов своего вида и содержали при различных комбинациях солености и температуры воды, соответствовавших наблюдаемым во время нерестового хода: при азовском (12-14 ‰, 24-25 °C), черноморском (18-19 ‰, 17-22 C°) течениях, а также при плавном снижении температуры воды и повышении солености. Созревания самок в искусственных условиях добивались с помощью инъецирования гормональными препаратами по методам, разработанным в ЮгНИРО [4, 6, 7, 9, 2].
Инъецирование самок лобана проводила Л. Г. Гнатченко, используя свежие и ацетонированные гипофизы своего вида. Икра сингиля и пиленгаса была получена нами совместно с Н. И. Куликовой, с 1993 г. икру пиленгаса получали совместно с А. Ф. Булли.
Результаты гормональной обработки оценивали по состоянию ооцитов в щуповых пробах после действия первой и последующих доз гормонального препарата [1].
Икру оплодотворяли «полусухим» способом и инкубировали в лотках и пластиковых бассейнах объемом от 50 до 400 л при плотности закладки 1 тыс. шт./л при температуре 16-23 °С.
На свежем материале измеряли диаметр овулировавшей икры и жировой капли (n = 25-50 для каждой самки), подсчитывали количество икринок в навеске, взвешенной на аналитических весах, и определяли среднюю массу одной икринки. Содержание влаги в икринках определяли высушиванием навески 0,5- 1,5 г при температуре 65 °C до постоянной массы. По массе высушенной навески определяли среднюю сухую массу икринки.
Для определения биохимического состава икру фиксировали в смеси Фолча. Содержание общих липидов, обезжиренного сухого вещества и влаги определяли методом Фолча в модификации Лапина и Черновой [8].
Икра каждой самки осеменялась спермой 3-4 самцов и инкубировалась в отдельных емкостях в соответствии с методиками [7, 2].
В табл. 1 приведены результаты экспериментов по созреванию самок лобана в разных условиях при одинаковой схеме введения гипофизарных гормонов.
Как видно из таблицы, во все варианты опыта были отобраны самки с ооцитами, близкими по исходному состоянию, т. е. достигшими дефинитивных размеров.
Однако результаты различались как по числу самок, отреагировавших созреванием на гормональные инъекции, так и по качеству икры.
В условиях, близких к условиям нагула в Азовском море, при солености 13- 14,5‰ и повышающейся температуре воды с 23 до 25,5 °С, созревание лобана заканчивается овуляцией икры лишь у части рыб – 5-10 %. Процент ее оплодотворения, как правило, низкий – 27-29 %.
Как показано нами ранее [5], содержание рыб в этих условиях более 3 суток приводит к утрате их способности реагировать на введение гипофизарных гормонов, быстрому ухудшению самочувствия производителей: потере чешуи, появлению на этих участках тела кровоподтеков и дегенеративным изменениям в ооцитах.
В то же время, в условиях постепенно повышающейся солености и понижающейся температуры, практически у всех рыб наблюдалась овуляция зрелой икры (табл. 1). При этом отмечалась тенденция повышения сухой массы овулировавшей икры по мере снижения температуры воды во время созревания производителей. Этот показатель, по-видимому, является одной из важнейших рыбоводных характеристик качества икры. Ранее нами [3] было показано, что величина сухой массы овулировавшей икры кефалей, представляющей собой в основном трофические компоненты (белок и липиды), за счет которых развиваются будущие эмбрион и личинка, положительно коррелирует с размером желточного мешка личинок и их выживаемостью до 15-суточного возраста.
Очевидно, сроки миграций, их продолжительность, а также структура популяций и вариабельность физиологического состояния производителей кефалей в период нерестовыхмиграций связаны, прежде всего, с особенностями преднерестового развития гонад, биологии видов и степени совершенства у них механизма адаптации к созреванию и размножению (воспроизводству) в водах низкой солености.
Так, период нереста у сингиля, в отличие от такового у лобана, очень короткий. Кроме того, нерест сингиля происходит осенью и осложняется частыми сменами погодных условий: штормами, изменением направления течений, нередко резким снижением температуры воды.
Нерестовые миграции из Азовского моря в Черное у сингиля начинаются обычно в середине августа и продолжаются около месяца. В начале хода у большинства самок гонады находятся на III стадии зрелости, ГСИ варьирует от 4 до 17,9 %, средний диаметр ооцитов – от 125 до 450 мкм. Среди мигрирующих рыб в этот период преобладают самцы в соотношении 4:1 и 8:1. Самки с гонадами IV стадии зрелости (размер клеток – 470-540 мкм) составляют не более 10 %.
В пик хода соотношение полов становится равным 1:1, а в конце миграции косяки состоят преимущественно из самок (3:1, 10:1). В разгар хода – в конце августа-первой декаде сентября и в его конце ооциты самок, как правило, находятся на незавершенной и завершенной IV стадии зрелости, ГСИ достигает 26,3 %. В этот момент иногда встречаются зрелые самки, у которых ГСИ составляет 60-70 %.
Таким образом, в конце августа-первой половине сентября через Керченский пролив мигрируют физиологически наиболее подготовленные к нересту производители сингиля. Однако даже в этот период лишь 30-50% самок имеют ооциты, достигшие дефинитивных размеров (> 500 мкм).
По нашим наблюдениям, период нерестового хода кефалей, когда в Керченском проливе происходит постепенное понижение температуры воды и повышение ее солености, наиболее благоприятен для созревания производителей. При
Таблица 1. Характеристика икры лобана, созревавшего в разных условиях

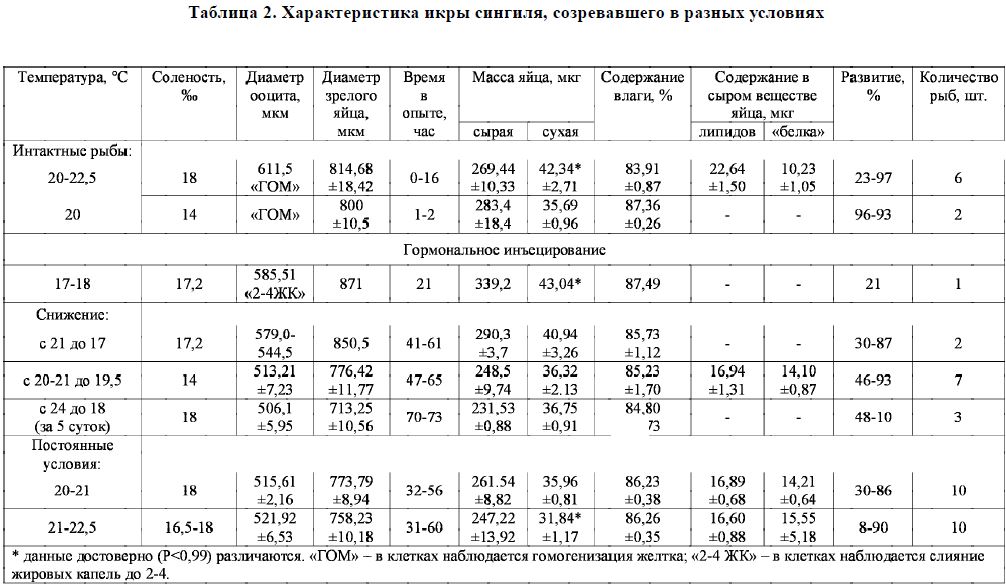
проведении рыбоводных работ в сериях опытов, в которых комбинации солености и температуры воды соответствовали наблюдаемым во время выхода этих рыб из теплых вод Азовского моря в более холодные воды Черного моря, нарушений созревания производителей не наблюдалось (табл. 2). В то же время, созревание рыб в 2007 г., при аномально высокой температуре воды 24-26 °С, проходило с нарушением и заканчивалось либо тотальной резорбцией, либо гибелью рыб. Лишь после снижения температуры воды в бассейнах замкнутой системы с 24 до 18 °С (в течение 5 суток) чувствительность к гормональным препаратам у самок повысилась, была получена зрелая икра (см. табл. 2).
Как видно из табл. 2, созревание самок сингиля при температуре 21-22,5 °С происходит за более короткий промежуток времени, чем при снижающейся температуре. Однако в последнем случае качество икры выше: по мере снижения температуры и повышения солености во время созревания производителей отмечается тенденция повышения величины основных рыбоводных показателей зрелых ооцитов. Средние показатели сухой массы икры в крайних вариантах различаются достоверно (Р < 0,95).
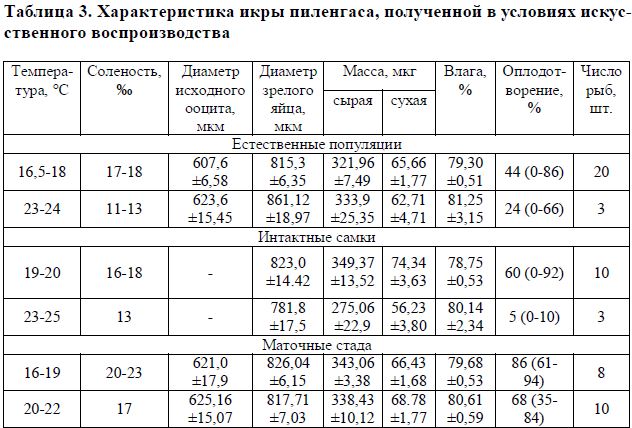
Сходная тенденция наблюдается также при сравнении показателей сухой массы икры «дикого» пиленгаса, созревавшего в разных условиях (табл. 3). В икре рыб маточных стад таких различий не обнаружено. Вероятно, это свидетельствует о том, что параметры среды, в которых проводились эксперименты по получению зрелой икры от рыб, выращенных в искусственных условиях, варьировали в пределах, близких к оптимальным для вида.
При температуре воды выше 24 °С у пиленгаса, как и у других кефалей, отмечаются нарушения в созревании ооцитов, появляются дегенеративные изменения. Содержание рыб в таких условиях уже через двое суток приводило к утрате их способности реагировать на введение гормональных препаратов, быстрому ухудшению самочувствия и тотальной резорбции половых клеток. По-видимому, введение гормональных препаратов при высокой температуре и низкой солености воды прерывает процесс нормального завершения вителлогенеза, что в итоге ведет к снижению сухой массы зрелого яйца.
Высокая температура блокирует не только созревание, но и процесс овуляции у кефалей. Оболочки неовулировавшей и невыметанной икры могут уплотняться, «спрессовываться» в твердые образования. Подобная картина отмечается и у самок черноморских кефалей (лобана и сингиля). Эти образования могут иметь значительные размеры, сохраняться в течение ряда лет и отражаться на степени развития гонад в последующие нерестовые сезоны.
В то же время, в условиях повышающейся солености (с 13-14 до 18-19‰) и плавно понижающейся температуры (с 22-20 до 18-16 °С) практически у всех рыб наблюдалась овуляция зрелой икры. Американские исследователи [10] также отмечали, что для осуществления нормального нереста кефалям необходимы более низкие температуры по сравнению с теми, при которых идет активный трофоплазматический рост.
Резкое снижение температуры воды приводит к резорбции ооцитов.
Таким образом, результаты наших экспериментов показывают, что нормальное завершение созревания ооцитов кефалей всех трех исследуемых видов происходит в температурном диапазоне – от 16 до 23 °C. При постепенном снижении температуры и повышении солености отмечается тенденция увеличения ряда
Таблица 2. Характеристика икры сингиля, созревавшего в разных условиях
важнейших рыбоводных характеристик овулировавшей икры: размеров, сырой и сухой массы, количества сухого обезжиренного вещества, а также ее оплодотворяемости. Созревание кефалей при 25-26°С в большинстве случаев сопровождается различными нарушениями, часто появляются признаки резорбции желтковых ооцитов. У рыб снижается чувствительность к гормональным препаратам, отмечаются покраснение плавников, потеря чешуи, а нередко и гибель. Содержание рыб во время экспериментов в замкнутых рециркуляционных системах, в которых температурные и соленостные параметры среды соответствуют оптимальным (17-22 °С и 17-20 ‰ – предпочтительно плавное снижение первых и повышение вторых в пределах указанных значений), значительно повышает эффективность рыбоводных работ, позволяет избегать негативных влияний внешней среды.
Следовательно, для нормального завершения вителлогенеза в яйцеклетках азово-черноморских кефалей, обеспечивающего повышение уровня чувствительности к гормональным препаратам и качества половых продуктов, необходимы регулируемые условия, которые могут быть созданы только в замкнутых системах и термостатированных помещениях.
Таблица 3. Характеристика икры пиленгаса, полученной в условиях искусственного воспроизводства
Литература
1. Апекин В. С., Вальтер Г. А., Гнатченко Л. Г. Изменение ооцитов при
созревании и получении зрелой икры с помощью гомопластических инъ-
екций у лобана (Mugil cephalus L.) // Труды ВНИРО. – М., 1976. – Т. 115. –
С. 13-23.
2. Биотехника искусственного воспроизводства кефалей (лобана, сингиля,
пиленгаса) с описанием схемы типового рыбопитомника. / Составители:
Куликова Н. И., Шекк П. В. – Керчь: ЮгНИРО, 1996. – 27 с.
Таблица 3. Характеристика икры пиленгаса, полученной в условиях искус-
ственного воспроизводства
3. Булли Л. И. Связь общего биохимического состава икры пиленгаса с ка-
чеством получаемого потомства // Труды ЮгНИРО. – Керчь: ЮгНИРО,
1996. – Том 42. – С. 221-224.
4. Вальтер Г. А. Особенности созревания самок сингиля (Mugil auratusRisso)
в зависимости от размера их ооцитов, а также дозы введенного гипофиза
сазана // Физиология морских рыб. – М.: Пишевая промышленность, 1980.
– С. 70-77.
5. Гнатченко Л. Г., Федулина В. Н., Булли Л. И. Определение опти-
мальных условий преднерестового содержания черноморского лобана
для перевода в нерестовое состояние // Тезисы докл.: VIII науч. конф.
по экологической физиологии и биохимии рыб. Т. 1. – Петрозаводск,
1992. – С. 69-70.
6. Инструкция по разведению кефали лобана / Составители: Аронович Т. М.,
Маслова О. Н., Лапина Н. М., Куликова Н. И., Гнатченко Л. Г., Демьяно-
ва Н. И., Куприянов В. С., Шершов С. В. – М.: ВНИРО, 1986. – 54 с.
7. Инструкция по разведению кефали-сингиля / Составители: КуликоваН. И.,
Демьянова Н. И., Хомутов С. М., Гнатченко Л. Г., Федулина В. Н., Се-
мик А. М., Куприянов В. С., Макухина Л. И., Писаревская И. И., Копей-
ка Н. В., Фитингов Е. М. – М.: ВНИРО, 1990. – 69 с.
8. Лапин В. И., Чернова Е. Г. Ометодике экстракции жира из сырых тканей
рыб // Вопросы ихтиологии. – 1970. – Т. 10. – Вып. 4. – С. 753-756.
9. Методические указания по разведению кефали-пиленгаса Mugil so-iuy
(Basilewsky) в водоемах юга Украины / Шекк П. В., Куликова Н. И., Феду-
лина В. Н., Яровенко А. В., Макухина Л. И., Булли А. Ф., Воля Е. Г. –
Киев: Укррыбхоз, 1993. – 19 с.
10. Kuo C.-M., Nash C. E., Shehadeh Z. H. The effects of temperature and
photoperiod on ovarian development in captive greymullet (Mugil cephalus L.)
// Aquaculture. – 1974 b. – V. 3. – Nо 1. – P. 25-43.
Л. И. Булли
ЮгНИРО