РЕОРЕАКЦИЯ И ФОРМИРОВАНИЕ ФЕНОТИПИЧЕСКИХ ГРУПП СЕГОЛЕТОК АТЛАНТИЧЕСКОГО ЛОСОСЯ
Экспериментально исследовали локомоторные показатели реореакции сеголеток атлантического лосося двух фенотипических групп, образовавшихся после расселения из близко расположенных нерестовых гнезд. Показано, что заселение микробиотопов, имеющих различные гидрологические характеристики, определяется реореакцией мальков. Часть из них проявляла повышенную способность сопротивляться потоку и заселяла микробиотопы с высокими скоростями течения. В дальнейшем это должно отразиться на формировании возрастных групп по срокам обитания в реке (от 2 до 4 лет) и на наступлении смолтификации.
Данные подводных наблюдений 20052007 гг. показали, что в прибрежной части порога Ареньгский основного русла р. Варзуги и в устье впадающего в него притока Ареньга ежегодно к IV декаде июня образуются скопле-
ния ранних сеголеток лосося. Происходит это в результате их расселения из нерестовых гнезд центральной части порога [Pavlov et al., 2010]. По возникшим в течение летнего периода различиям в размерно-весовых и биохимических показателях (энергетического обмена и липидного статуса) было доказано формирование из
0
этих скоплений устойчивых фенотипических групп [Павлов и др., 2007а,б, 2008].
Известно, что основной врожденной поведенческой реакцией рыб, обитающих в потоке, является реореакция, она компенсирует снос рыб против течения и способствует их удержанию в районе обитания [Павлов, 1979]. В онтогенезе молоди рыб существенные изменения показателей происходят на первых этапах развития, так как интенсивно меняется морфология тела, развиваются локомоторные органы, органы чувств и плавательная способность [Васнецов, 1953; Алеев, 1963; Павлов, 1979, 1986]. Для реореакции это сопровождается снижением пороговых и увеличением критических скоростей течения. Реореакция может служить мерой отношения к потоку и быть ключом к раскрытию адаптивных механизмов, реализуемых в поведении рыб. Она влияет на закономерности пространственного расселения рыб и соответственно на возникновение фенотипических групп сеголеток лосося.
В связи с этим задача работы заключалась в изучении роли реореакции при расселении сеголеток лосося из нерестовых гнезд и формировании фенотипических групп.
Материалы и методы
Наблюдения за расселением сеголеток из нерестовых гнезд и образованием прибрежных и устьевых групп проводили в 2005-2007 гг. с помощью подводного метода, позволяющего получить достоверные данные [Keenleyside, 1962; Wankowski, Thorpe, 1979; Павлов и др., 1981]. Используя аппарат электролова (Fa-2, Норвегия), сеголеток (0+) собирали с площадей микробиотопов в прибрежной части порога главного русла р. Варзуги и в устье притока Ареньга (66°32'43” с.ш., 36°12'06” в.д.). Мальков (в каждой пробе 30 экз.) отлавливали с момента образования устойчивых групп в послед-
ней декаде июня и повторно в первую декаду августа, выдерживая в русловых садках в течение суток до начала эксперимента.
Исследовали локомоторную компоненту реореакции сеголеток лосося. Эксперименты проводили в специально сконструированной гидродинамической установке (рис. 1). Экспериментальная камера представляет собой стеклянную трубку диаметром 20 мм и длиной 1 м, в головной части которой находятся успокоители потока. Скорость течения внутри рабочей камеры изменяли в пределах 0-0,8 м/с при помощи двух кранов, установленных перед трубкой. В рабочую камеру, предварительно наполненную водой, через разъем сливного патрубка пересаживали по одному мальку из исследуемой фенотипической группы.
У каждого малька измеряли три показателя реореакции с трехразовой повторностью. Пороговая скорость течения (Упор.) для реореакции регистрировалась при медленном (30-40 с) увеличении скорости течения от нулевого значения до момента, когда малек начинал проявлять двигательную реакцию в виде нистагма глаз, изгиба плавников или хвостовой части тела. Показатель Уа|СТ характеризовал достижение (за 15-20 с) такой скорости течения, при которой ориентированный против течения малек, находясь в тактильном контакте с дном, переставал удерживаться только за счет прижимного давления потока на плавники и включал активную локомоцию. Критическая скорость (Укрит) течения (выставлялась за 10 с) отмечалась при прекращении мальком сопротивления наибольшей скорости течения и сбросе его на защитную сетку. После эксперимента мальков взвешивали и измеряли длину.
Скорость течения в экспериментальной камере находили, измеряя объем воды (^ в мерном сосуде и время (1) его наполнения из сливного патрубка. Объем воды в мерном сосуде в опытах изменялся в пределах от 50 до 1000 см3.
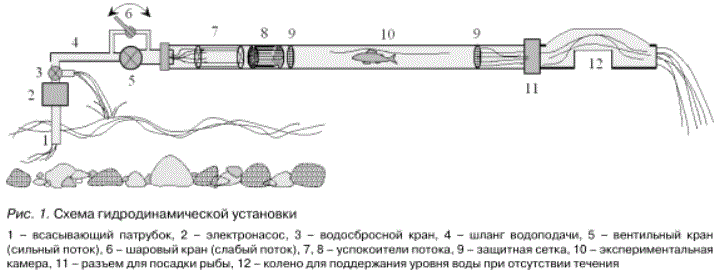
Рис. 1. Схема гидродинамической установки
1 - всасывающий патрубок, 2 - электронасос, 3 - водосбросной кран, 4 - шланг водоподачи, 5 - вентильный кран (сильный поток), 6 - шаровый кран (слабый поток), 7, 8 - успокоители потока, 9 - защитная сетка, 10 - экспериментальная камера, 11 - разъем для посадки рыбы, 12 - колено для поддержания уровня воды при отсутствии течения
По этим величинам рассчитывали расход воды в трубе О = W/t, а затем вычисляли среднюю скорость течения - V = О/Э, где Э - площадь поперечного сечения трубы, Э = 3,14 см2.
Результаты и обсуждение
Образование фенотипических групп сеголеток лосося. Нерестовые гнезда атлантического лосося с вылупившимися эмбрионами располагались в центральной части главного русла р. Варзуги. С помощью подводных наблюдений установлено, что в течение 2-3 суток при подъеме температуры воды от 9 до
12-13°С личинки (обычно к 12-16 июня) появляются в поверхностном слое галечного грунта (рис. 2 а). В области расположения нерестовых гнезд плотность личинок достигала
13-17 экз./м2. Затем в течение пяти суток происходило их массовое активно-пассивное расселение, в основном по направлению речного
потока. В этот период личинки полностью переключались на экзогенное питание, у них закладывался чешуйный покров и завершался переход на стадию малька. Плотность распределения в шлейфе расселения снижалась до 3, реже 7 экз./м2 (рис. 2 б). На седьмые сутки мальков обнаруживали на границе порога с плесом, одновременно прекращалось их расселение по течению (рис. 2 в). Плотность распределения колебалась в тех же пределах, однако на восьмой-девятый день преобладающим становилось активное, против потока перераспределение из случайно заселенных микробиотопов вдоль границы порога к прибрежным участкам главного русла или в устье притока Ареньга (рис. 2 г, д). Здесь, в прибрежье и устье ежегодно образовывались фенотипические группы (рис. 2 е), из которых мальков отлавливали для эксперимента (участки I и II). Плотность распределения в этих группах к концу июня составляла 2-12 экз./м2.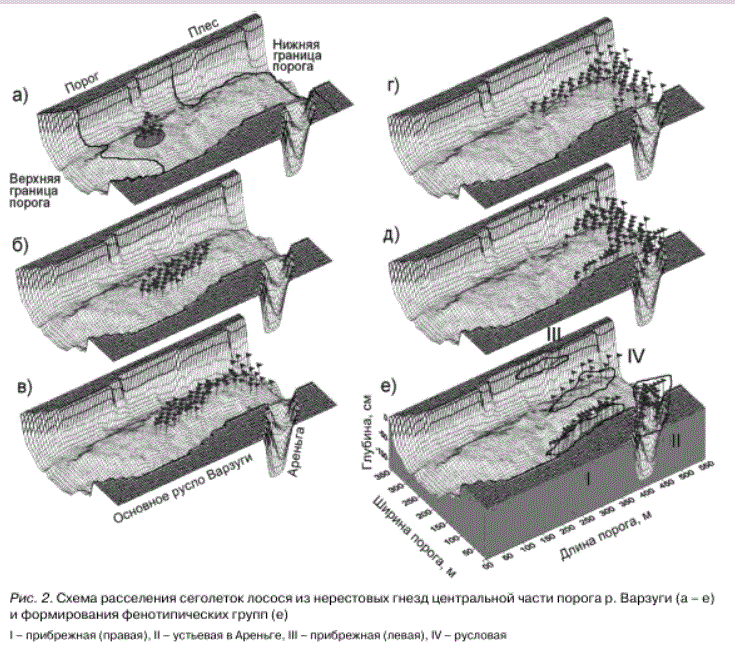
Рис. 2. Схема расселения сеголеток лосося из нерестовых гнезд центральной части порога р. Варзуги (а - е) и формирования фенотипических групп (е)
I - прибрежная (правая), II - устьевая в Ареньге, III - прибрежная (левая), IV - русловая
Сравнение микробиотопов показало, что в притоке (участок II) грунт крупнее. Он состоит на 35-45 % из мелких (5-10 см) и на 40-50 % из средних размеров валунов (11-25 см), содержание гальки (1,0-4,9 см) не превышало 15 %. На таком грунте хорошо представлены нитчатые водоросли и обрастания мха, способствующие развитию кормовой базы - зообентоса беспозвоночных организмов [Шустов, 1983]. Фракционный состав грунта прибрежного правого участка (I), где отлавливали мальков, напротив, состоял преимущественно из гальки (до 65-80 %) и мелких валунов (15-30 %), а также содержал незначительное (около 5 %) количество валунов средних размеров. Этот грунт почти не имел обрастаний. Оценка варьирующих глубин в целом не выявила существенных различий по микробиотопам, однако средняя поверхностная скорость течения в притоке Ареньга (0,97 м/с), кроме периода малой воды в августе, была достоверно выше за счет большего уклона русла, чем в прибрежье Варзуги (0,58 м/с).
Таким образом, участок обитания в притоке за счет более крупных фракций грунта и повышенных скоростей течения обеспечен более разнообразными микростациями для молоди лосося. Это также подтверждалось плотностью распределения: 7-12 экз./100 м2 в притоке и 2-5 экз./100 м2 в прибрежье. Далее экспериментально исследовали образовавшиеся две фенотипические группы сеголеток лосося на различия в локомоторных показателях реореакции.
Локомоторные показатели реореакции фенотипических групп сеголеток лосося. Экспериментально установлено, что минимальная скорость течения (Упор), стимулирующая двигательную реакцию в виде нистагма глаз, изгиба плавников, достоверно не различается между особями обоих групп как в июне, так и в августе (табл.). Можно предположить, что проявление реореакции на пороговую скорость течения, или чувствительность к потоку, изначально одинакова у сеголеток лосося и различия, как показано [Veselov et al., 1998; Веселов и др., 2001], проявляются лишь в возрасте 1+ и старше. Следовательно, показатель V^ менее существен для избрания будущего микробиотопа обитания, чем способность лавировать и физически противостоять неоднородному турбулентному потоку при совершении пищевых или оборонительных бросков и перемещении в придонном пространстве.
В связи с этим при усилении потока исследовали более значимые для активного освоения подвижного пространства обитания показатели V и V
акт. крит.
Показатель V3CT характеризует достижение такой скорости течения, при которой малек, находясь в тактильном контакте с дном, перестает
удерживаться только за счет прижимного давления потока на развернутые им грудные и брюшные плавники. Происходит смещение его относительно неподвижных ориентиров, что и стимулирует включение локомоции, обеспечивающей возврат и устойчивое поддержание исходной позиции. Установлено, что в июне показатель Уакг, измеренный у мальков прибрежной группы основного русла р. Варзуги, достоверно меньше, чем у мальков из устья притока. К августу эти различия усиливаются (см. табл.). Это свидетельствует о раннем включении локомо-ции прибрежными сеголетками и, следовательно, совершаемыми ими повышенными энергетическими тратами в одинаковых с устьевыми мальками условиях потока.
Другой локомоторный показатель реореак-
ции - V
характеризует краткосрочно вы-
держиваемую максимальную для малька скорость потока. Его значения отражают способность рыбы совершать пищевые или оборонительные броски из спокойных по течению придонных слоев в толщу воды, характеризующуюся стремительным потоком. Например, на таких участках порогов, где поверхностная скорость течения около 1,5-2 м/с, что не редкость для лососевых нерестовых рек [Веселов, Ка-люжин, 2001; Калюжин и др., 2009]. Экспериментально нами выявлены достоверные различия по этому показателю между обеими группами в пользу устьевых мальков (II), причем они возрастают в летний период (см. табл.).
Анализируя оба показателя (Уакг и Укрит), можно сказать, что в динамичных по структуре потока речных условиях обитания для устьевых сеголеток с развитием реореакции и локомоторных способностей повышается доступность по градиенту скоростей течения разнообразных микробиотопов и микростаций [Веселов, Калюжин, 2001]. Это в свою очередь связано с широтой вариации кормовых условий [Барышев, Веселов, 2003; Барышев, 2004], определяющих темп роста [Шустов, Смирнов, 1978]. Например, исследования бентоса в бассейне р. Варзуги показали, что в среднем количество амфибиотиче-ских насекомых существенно больше в притоках, чем в главном русле [Барышев, Веселов, 2003; Барышев, 2004], поэтому к августу среди образовавшихся групп проявились четкие размерно-весовые различия (см. табл.).
Во многих притоках нерестовых рек, имеющих сложную гидрографическую структуру, складывается более благоприятное для сеголеток лосося и развития кормовых организмов сочетание рельефа и гидрологических факторов (глубина, скорость течения, фракционный состав грунта). В июне - начале июля в прибрежных микробиотопах Варзуги поверхностная скорость течения была ниже, чем в устье
Сравнительная характеристика показателей реореакции для прибрежной и устьевой фенотипических групп сеголеток лосося р. Варзуги в июне и августе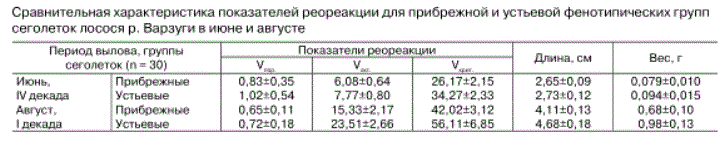 Ареньги. Установлено, что интенсивность заселения мальками притоков выше, чем их продвижение вдоль берега главного русла. Например, за 5-6 суток они перемещались от устья вверх по притоку на 250-300 м, а обитающие в прибрежье сеголетки - вдоль берега против течения всего на 100-150 м. Существенно и то, что разница дневных и ночных температур в притоках больше (5-7°С), чем в главном русле (3-5°С) [Павлов и др., 2008]. Это также может оказывать положительное влияние на рост молоди и накопление фенотипических различий [Сущеня, 1978; Константинов и др., 1987, 1989; Веселов, Калюжин, 2001], отражающихся на проявлении «силовых» (V3CT и Укрит) показателей локомоторной компоненты реореакции и способности осваивать микробиотопы с высокими скоростями течения.
Ареньги. Установлено, что интенсивность заселения мальками притоков выше, чем их продвижение вдоль берега главного русла. Например, за 5-6 суток они перемещались от устья вверх по притоку на 250-300 м, а обитающие в прибрежье сеголетки - вдоль берега против течения всего на 100-150 м. Существенно и то, что разница дневных и ночных температур в притоках больше (5-7°С), чем в главном русле (3-5°С) [Павлов и др., 2008]. Это также может оказывать положительное влияние на рост молоди и накопление фенотипических различий [Сущеня, 1978; Константинов и др., 1987, 1989; Веселов, Калюжин, 2001], отражающихся на проявлении «силовых» (V3CT и Укрит) показателей локомоторной компоненты реореакции и способности осваивать микробиотопы с высокими скоростями течения.
Таким образом, заселение микробиотопов, имеющих различные гидрологические характеристики, определяется реореакцией мальков. Часть из них проявляла повышенную способность сопротивляться потоку и заселила микробиотопы с высокими скоростями течения. У молоди лососевых (Salmonidae), как и у ряда других рыб, существует дифференциация на группы по морфологическим и физиолого-био-химическим показателям [Thorpe, 1977; Павлов и др., 2007б и др.]. Одна из предпосылок образования групп, по-видимому, связана с изначальной разнокачественностью икры по биохимическим показателям, например по липидному статусу [Нефедова и др., 2010]. Известно также, что икринки лососевых рыб, и в частности атлантического лосося, отличаются друг от друга по размерам, массе и объему запасенных веществ [Казаков, 1982]. Эта разно-качественность икры в последующем сказывается на интенсивности роста и развитии ранней молоди. Она влияет и на результаты первичного расселения сеголеток атлантического лосося из нерестовых гнезд [Веселов, Калюжин, 2001]. Можно предположить, что это определяет неодинаковые стартовые возможности расселения личинок в разные по условиям обитания микробиотопы и в значительной степени влияет на проявление такого интегрального показателя, как реореакция. Внутрипопу-ляционная неоднородность формируется под влиянием условий окружающей среды и встре-
чается даже у потомства одной пары производителей [Павлов и др., 2007а]. Ее адаптивное значение заключается в повышении выживаемости молоди [Павлов и др., 2010].
Заключение
Выявленные по показателям реореакции устойчивые различия сравниваемых групп сеголеток, которые возникли сразу после расселения из одних и тех же нерестовых гнезд, по-видимому, свидетельствуют о первичном влиянии разнокачественности эмбрионов на формирование различий по показателям реореак-ции. Наличие фенотипических групп у сеголеток лосося, по нашему мнению, играет значительную роль в дальнейшей дифференциации молоди лосося, поэтому проявившиеся различия реореакции можно рассматривать как одну из предпосылок возникновения фенотипических групп. В дальнейшем эти различия отразятся на выживаемости в зимний период, темпе роста, сроках смолтификации и миграции на нагул молоди в море. Неоднородные условия среды обитания, как и начальная разнока-чественность эмбрионов, вероятно, и определяют формирование сложной по числу прожитых лет в реке и море возрастной структуры атлантического лосося в крупных реках, например, Варзуге на Кольском полуострове [Казаков и др., 1992; Лысенко, Берестовский, 1999; Зубченко и др., 2002].
Работа выполнена при поддержке фонда РФФИ (грант № 11-04-00686_а).
А. Е. Веселов1, Д. С. Павлов2, М. А. Скоробогатов2, Д. А. Ефремов1, К. Ю. Потапов3
1 Институт биологии Карельского научного центра РАН
2 Институт проблем экологии и эволюции им. А. Н. Северцова РАН
3 Петрозаводский государственный университет
Смотрите также