МИГРАЦИОННОЕ ПОВЕДЕНИЕ ЛОСОСЯ И КУМЖИ В СРЕДНЕМ ТЕЧЕНИИ Р. КЮМИЙОКИ, ЮЖНАЯ ФИНЛЯНДИЯ (ПО ДАННЫМ БИОТЕЛЕМЕТРИЧЕСКОГО ПРОСЛЕЖИВАНИЯ)
Поскольку большинство нерестовых рек Финляндии зарегулировано, численность популяций анадромных рыб в последнее время поддерживается в основном за счет выпуска молоди искусственного воспроизводства в нижнем течении рек. В то же время на зарегулированных участках рек существуют потенциальные нерестилища лососевых рыб.
В историческом прошлом р. Кюмийоки служила местом размножения значительного числа анадромных видов (лосося, кумжи, а также сига). Природные популяции этих видов исчезли в результате зарегулирования стока и загрязнения воды в этой реке. Начиная с 80-х годов, интенсивное искусственное воспроизводство и выпуск молоди рыб в эстуарии реки создали основу успешного прибрежного рыболовства (Saura 1989; Saura, Mikkola 1990). Восстановления естественного воспроизводства анадромных рыб не произошло, так как плотины препятствуют нерестовой миграции производителей. Пересадка мигрирующих производителей ценных видов рыб, отловленных в нижних бьефах гидроузлов, по-видимому, может обеспечить естественный нерест на сохранившихся нерестилищах и получение жизнестойкой молоди рыб.
С целью выявления возможности использования производителями лосося Salmo salar L. и кумжи Salmo trutta L. сохранившихся нерестилищ в бассейне р. Кюмийоки проведены исследования поведения этих видов во время нерестовой миграции осенью 1992 и 1993 гг. Для изучения перемещений производителей лососевых применяли метод ультразвуковой биотелеметрии. Данная работа выполнена при поддержке финских коллег из НИИ охотничьего и рыбного хозяйства и Рыбохозяйственного отделения Кыми Финляндии: Ниеми А., Фриман Е., Пайваринта П., Вяярянен П., а также при активном участии Л.К. Малинина и В.Д. Линника — сотрудников Института биологии внутренних вод РАН.
Мигрирующие производители лосося и кумжи, подошедшие в нижний бьеф плотины Ахвенкоски (1992 г.) и Койвокоски (1993 г.), были отловлены, а затем выпущены выше плотины на разном удалении от нее в местах с разным гидрологическим режимом (рис. 1). Всего было помечено ультразвуковыми передатчиками моделей АРМ-30-15-1 и АРМ-50-15-1 (Саранчов 1983) 41 особь, в том числе производителей атлантического лосося — 27 (в 1992 г. — 15 , в 1993 — 12), производителей кумжи — 14 (в 1992 г. — 9, в 1993 г. — 5).
Ультразвуковые передатчики крепили наружно сбоку у основания спинного плавника по методике описанной Греем и Хайнесом (Gray, Haynes 1979). Перед мечением рыбу анестезировали раствором MS-222 (1992 г.). В 1993 г. в качестве анестетика для двух лососей применяли раствор бензокаина. Остальных рыб метили без анестезии, следуя рекомендациям Мохуса и Холланда (Mohus, Holland 1983). Мечение проводили в воде, постоянно насыщаемой кислородом воздуха. Рыб закрывали плотной тканью, прикрывая им глаза. Пеленгацию рыб осуществляли с помощью оригинальной отечественной аппаратуры двумя методами: триангуляции и наезда. Регистрацию местоположения меченых рыб проводили с мотолодок и с берега. На одной из лодок ежедневно объезжали места на участках последней регистрации особей.
В отдельных случаях, когда меченые рыбы проявляли повышенную двигательную активность, за ними проводили более детальные наблюдения, регистрируя местоположение рыб каждые 10-20 минут. За период исследования температура воды в реке снизилась с 16.0° С до 12.6° С. Скорость течения воды (по результатам замеров гидрологической вертушкой) на различных участках района исследования изменялась в пределах 20-156 см/сек. Данные наносили на планшетные карты 1:10000, по которым, затем рассчитывали скорости и направление движения.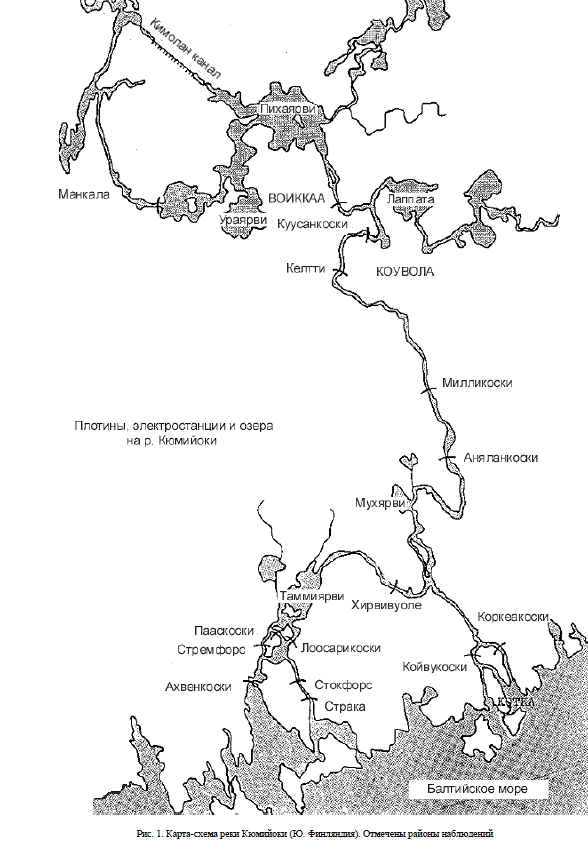
Рис. 1. Карта-схема реки Кюмийоки (Ю. Финляндия). Отмечены районы наблюдений 110
Atlantic salmon: biology, conservation and restoration, 2003
Зависимость скоростей и направления плавания меченых рыб от их пола, длины, длительности наблюдений, промежутки времени между точками наблюдений (elapsed time between observations) оценивали двумя способами. Во-первых, применяли Kruskal-Wallis тест, как непараметрический аналог одномерного анализа вариант. Ранжированный коэффициент корреляции Спирмена был подсчитан для вычисления отношения между скоростями движения, биологическими характеристиками меченых рыб и т.п. Из-за трудности оценки реальных скоростей плавания меченых рыб мы использовали в основном скорости плавания относительно ориентиров береговой линии. Естественно, что реальные скорости движения меченых рыб были более высокими, чем скорости движения относительно береговой линии, так как в дополнение к скорости движения самой рыбы добавляется скорость течения воды. Для расчета значительности различий скоростей плавания мы использовали тест Манн-Витни (Mann-Whitney, U test).
При анализе скоростей движения рыб в первую очередь выявляли статистический тип распределения исследуемых параметров. Исходя из него, выбирали статистические процедуры, оптимальные для анализа данных. Так, при статистически достоверных отличиях распределения варьирующих величин от закона нормального распределения, применяли непараметрические методы обработки. Например, для характеристики скоростей плавания использовали медиану, нижний и верхний квартили; для характеристики изменений скоростей плавания рыб в разные сутки наблюдений использовали multiple box-and-whisker (Tukey 1977), при этом для каждой подгруппы определяли медиану (горизонтальная черта в box), верхний и нижний квартили (границы box), range (vertical lines), flagged data (situated more then 1.5 interquartile intervals from median).
Сопоставление результатов выпуска рыб в трех разных участках (как в1992, так и в1993 гг.) показало, что наименее пригодна в качестве места выпуска верхняя камера рыбохода. Выпуск меченых рыб в нее приводил к большим затратам энергии меченых рыб на сопротивление потоку воды в рыбоходе. Это не позволяло рыбам восстановить силы после стресса, вызванного мечением и хэндлингом. В результате, из двух рыб (лосося и кумжи), высаженных в верхнюю камеру рыбохода, крупный самец лосося, после того, как скорость течения воды в рыбоходе была уменьшена, вышел из верхней камеры рыбохода в приплотинную зону верхнего бьефа регуляционной плотины, и уже через 20 минут скатился вниз по течению через плотину, а кумжа погибла. Таким образом, пример этих рыб показывает, что даже для таких рыб с высокой плавательной способностью, как лосось и кумжа, выпуск меченых особей в камеры рыбоходов с высокими скоростями течения не желателен.
Значительно более приемлемыми для обеспечения хороших условий для отдыха меченых особей сразу после их выпуска оказались два других места выпуска – в 50 м и 700 м выше регуляционной плотины ГЭС Койвокоски, где скорости течения не превышали 30 см/сек. Меченые особи, выпущенные на этих участках, продемонстрировали обычно наблюдавшееся нами поведение лососевых: сразу после выпуска – медленное перемещение вниз по течению с последующими поисковыми перемещениями, или остановка в районе выпуска на нескольких часов.
Характерные стереотипы поведения меченых рыб приведены на рис. 2. Среди исследованных рыб особи с разной реореакцией были представлены в различных долях (табл. 1).
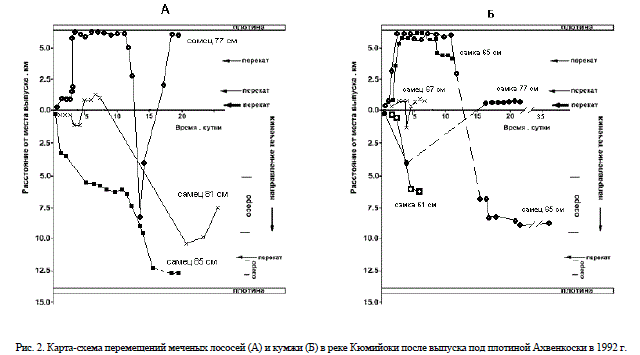
Рис. 2. Карта-схема перемещений меченых лососей (А) и кумжи (Б) в реке Кюмийоки после выпуска под плотиной Ахвенкоски в 1992 г.
Таблица 1.
Соотношение меченых рыб в группах за весь период наблюдения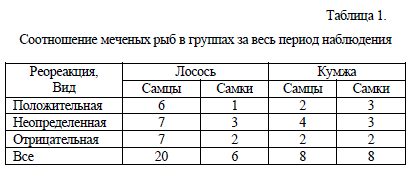
Около 1/3 части лососей продемонстрировали положительный реотаксис. К этой группе рыб принадлежали 4 самца. После выпуска эти рыбы проявили кратковременный скат вниз по течению, затем в течение нескольких часов отстаивались на одном месте, после чего начинали подъем против течения либо в день выпуска, либо к концу вторых суток после выпуска. У лососей первой группы была выраженная тенденция к сумеречно-ночной двигательной активности.
В поведении второй группы лососей доминировали маломасштабные поисковые перемещения сравнительно недалеко от точки выпуска. Для этих рыб были характерны как довольно быстрый скат с последующей длительной остановкой в приплотинном участке верхнего бьефа, так и выраженные поисковые перемещения различного масштаба в районе выпуска с тенденцией к перемещению вниз по течению. Таким образом, и для лососей второй группы был характерен достаточно широкий спектр двигательных реакций.
Более стереотипным было поведение третьей группы лососей, скатывавшихся вниз по течению, первая особь скатилась через плотину вскоре после выхода из верхней камеры рыбохода у регулировочной плотины, а вторая скатилась вниз через плотину на третьи сутки после выпуска.
Меченые кумжи проявили положительный реотаксис. Подъем рыб вверх происходил в 1-2-ые сутки после выпуска. Для них были не характерны выраженные поисковые перемещения. Миграция кумж вверх происходила как в темное, так и в светлое время суток. Самка кумжи, относившаяся ко второй группе рыб по реореакции, в первые часы после выпуска скатилась вниз по течению к плотине ГЭС Койвокоски и находилась вблизи плотины 10 суток
Вариационные ряды скоростей плавания как лососевых первых двух групп, так и кумж первой группы имели четко выраженные асимметрию и эксцесс. Замечено, что лососи и кумжа, активно мигрирующие против течения, имели несколько различный характер варьирования скоростей плавания. У кумжи доля высоких скоростей плавания была больше, чем у лосося. Статистические показатели скоростей плавания меченых рыб приведены в табл. 2. Гистограммы скоростей плавания меченых лососевых, статистически достоверно отличаются от нормального распределения.
При сопоставлении скоростей плавания меченых рыб в 1992 и в 1993 гг. выявлено, что скорости плавания лососей первой группы достоверно не отличались (U=1.67, P<0.1), скорости плавания лососей второй группы в 1992 г. были статистически достоверно выше, чем в 1993 г. (U=4.38, P<0.00001), для кумжи первой группы скорости плавания в 1992 г. были достоверно ниже (U=2.97, P<0.003).
Приведенные выше значения скоростей плавания рассчитаны для всего периода наблюдений, т.е. сюда вошли данные наблюдений, когда рыбы не проявляли активных перемещений, отстаиваясь после преодоления порогов или находясь в приплотинной зоне нижнего бьефа ГЭС Ахвиокоски. Чтобы избавиться от искажающего влияния указанных отсчетов (большое число наблюдений), мы сделали попытку оценить максимально возможные скорости миграции меченых нами рыб в начальной фазе движения вверх по течению (табл. 3).
Приведенные в табл. 3 значения скоростей плавания в начальный момент миграции значительно расходятся с таковыми для лососей естественной популяции из шотландской реки Dee. Лососи из этой популяции имели более высокую скорость движения вверх по реке (1.68-21.99 км/день, а у нас – 9.49 км/день) и более длительный период подъема по реке с высокой скоростью (0.84--17.11 дней, а у нас – 69 дней) (Hawkins & Smith 1986) по сравнению с нашими подопытными рыбами. По данным указанных авторов, практически все рыбы (10 лососей), меченые передатчиками, быстро преодолели эстуарий и нижнюю часть реки, а затем скорость их движения замедлялась. По сравнению с полученными в нашей работе данными схемы подъема рыб, приведенные Hawkins, Smith (1986) были значительно единообразнее.
По нашим наблюдениям (test Kruskal-Wallis) у лососей первой группы скорость плавания зависела от длительности прослеживания и от направления перемещения рыб; у лососей второй группы выявлена связь скоростей плавания с длиной рыб, временем определения местоположения и направлением их перемещений (табл. 4).
По данным корреляционного анализа видно, что скорость плавания рыб относительно берега у всех приведенных групп достоверно коррелирует со временем между регистрациями местоположения рыб и направлением движения (табл. 5).
Таблица 2.
Статистические характеристики движения меченых рыб в 1992-1993 гг.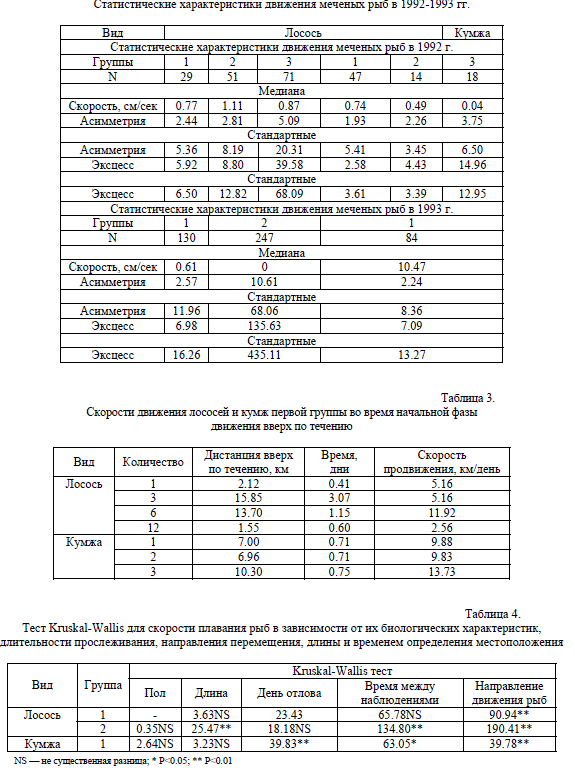
Таблица 5.
Spearman коэффициенты корреляции между направлением, скоростью плавания, биологическими характеристиками и временем между регистрациями 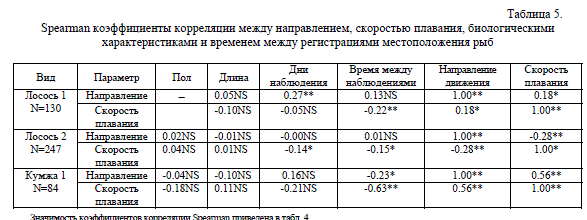 Значимость коэффициентов корреляции Spearman приведена в табл. 4.
Значимость коэффициентов корреляции Spearman приведена в табл. 4.
У группы особей, не проявивших положительного реотаксиса, скорость плавания также отрицательно коррелировала с длительностью наблюдений, т.е. по мере наблюдений за этими рыбами их скорости плавания падали. Следует отметить, что для обеих групп лососей и группы кумжи с положительной реореакцией была характерна отрицательная связь скорости плавания со временем между наблюдениями (т.е. наивысшие расчетные скорости плавания были отмечены при минимальных значениях времени между наблюдениями местоположения рыб). У групп лососей и кумж с выраженной положительной реореакцией был одинаков характер связи скоростей плавания с некоторыми параметрами: отрицательный со временем между наблюдениями и положительный с направлением движения (т.е. при движении рыб против течения их скорость плавания относительно берега была выше, чем при движении вниз по течению). У лососей обеих групп выявлена достоверная отрицательная связь скоростей плавания и длительности наблюдений. Аналогичная зависимость была обнаружена у лосося р. Пенобскот (Power, McCleave 1980). По-видимому, в этом проявляется общая закономерность снижения скоростей плавания мигрирующих производителей анадромных рыб по мере их приближения к нерестилищам.
Интересно сравнить направление движения рыб различных групп в течение всего периода прослеживания. У лососей и кумжи первой группы в начальный период наблюдений немного преобладали перемещения против течения и остановки, к концу периода наблюдений отсутствовали перемещения вниз-вверх по течению, рыбы предпочитали находиться на определенных участках. У лососей второй группы в течение всего периода наблюдений перемещения вверх-вниз по течению и остановки были равновероятными.
Скорее всего, значительное разнообразие трасс движения половозрелых особей лосося, полученных от искусственного воспроизводства, является следствием неадаптированности механизмов ориентации вселяемых рыб к конкретным условиям водоема. Подтверждение этому положению можно найти в работе Power, McCleav, (1980), исследовавших трассы перемещений половозрелых атлантических лососей, вселенных на стадии смолтов на различных участках реки Пенобскот (Maine, USA). Движение этих рыб происходило не только по реке вверх, но часто перемежалось продолжительными остановками и сплыванием вниз по течению. Часть рыб не проявила положительного реотаксиса. Для схем движения указанных рыб была характерна такая же высокая степень разнообразия, как и для лососей, прослеженных нами в р. Кюмийоки. О высокой степени изменчивости миграционного поведения производителей лосося, полученных от искусственного воспроизводства, свидетельствует тот факт, что при прослеживании перемещений лососей в р. Пенобскот 20 меченных радиопередатчиками особей прошли через рыбоходы плотин в верхний бьеф, а затем скатились вниз по течению через плотины, и лишь 4 особи не скатились, пройдя этот путь (Power, McCleave 1980). Таким образом, стереотипы поведения производителей лососевых, полученных от искусственного воспроизводства, характеризуются высокой степенью неустойчивости и большой долей вероятностного компонента. Следует отметить, что меченные нами производители кумжи имели значительно более устойчивый стереотип миграционного поведения, чем лососи. Это тем более удивительно, что в системе р. Кюмийоки при зарыблении смолтов лосося выпускали как в средней, так и в нижней зонах речной системы, а смолтов кумжи выпускали в эстуарии.
Для мигрирующих анадромных производителей природных мигрирующих популяций (of nature spawning populations) более свойственны устойчивые стереотипы миграционного поведения — направленное движение к нерестилищам с отсутствием блужданий (Hawkins, Smith 1986; Heggberget 1988). Существенные отличия в поведении производителей атлантических лососей, выросших от диких и смолтифицирующихся в эстуарии farmed особей, отмечены и норвежскими исследователями. Для производителей от искусственного и естественного воспроизводств было характерно одновременное появление вблизи эстуария нерестовой реки, но wild fishes заходили в реку раньше и уходили позже, чем рыбы от искусственного воспроизводства. Последние имели более высокий уровень двигательной активности, сильнее травмировались в реке, и 13.5 %
114
Atlantic salmon: biology, conservation and restoration, 2003
самок и 36.7 % самцов уходили из реки в море не отнерестившись. Из диких производителей в реке нерестились все самки и 96.2 % самцов (Johnson et al. 1990).
Полученные данные выявили интересные стороны миграционного поведения рыб, полученных от искусственного воспроизводства, выпущенных в виде молоди в верхней части эстуария реки. После нагула в море созревшие рыбы вернулись в верхнюю часть эстуария. При пересадке производителей этих рыб в верхний бьеф плотины, фактически на незнакомый для них участок реки, рыбы демонстрировали достаточно разнообразное поведение.
У лососей и кумжи по стереотипам поведения среди меченых рыб можно выделить три группы особей. Во-первых, рыбы, активно реагирующие на поток и стремящиеся двигаться вверх против течения до непреодолимой преграды. Часть этих рыб продолжала оставаться вблизи препятствия, одна особь после ската вновь поднялась к плотине. Во-вторых, особи, совершающие поисковые перемещения вверх и вниз по течению. В-третьих, рыбы, отрицательно реагирующие на течение и, как правило, скатывающиеся вниз по течению. У лососей и кумжи, активно мигрировавших против течения, варьирование скоростей плавания имело различный характер — у лосося доля высоких скоростей плавания была меньше, чем у кумжи.
Таким образом, стереотипы поведения производителей лососевых, полученных от искусственного воспроизводства, характеризуются высокой степенью неустойчивости и большой долей вероятностного компонента. Следует отметить, что меченные нами производители кумжи имели значительно более устойчивый стереотип миграционного поведения, чем лососи. Как уже указывалось, это тем более удивительно, что в системе р. Кюмийоки при зарыблении смолтов лосося выпускали как в средней, так и в нижней зонах речной системы, а смолтов кумжи выпускали в эстуарии.
Таким образом, анализ полученных и литературных данных показал, что значения скоростей плавания в период миграции у производителей (искусственное воспроизводство) отличаются от таковых у особей из естественных популяций лососевых. Схемы подъема рыб из естественных популяций более единообразны, чем полученные в наших наблюдениях. Скорее всего, значительное разнообразие трасс движения половозрелых лососей, полученных от искусственного воспроизводства, является следствием неадаптированности механизмов ориентации вселяемых рыб к конкретным условиям водоема.
Полученные данные, несомненно, важны для рыбохозяйственных целей и представляют интерес для оценки адаптационных возможностей лососей искусственного и естественного воспроизводства. При попытках реинтродукции лососевых в водоемы, где эти рыбы ранее обитали, первые две группы рыб могут создать ядро нерестовой популяции. Возможно, увеличение доли производителей, активно разыскивающих нерестилища, за счет массовых выпусков молоди конспецифичных лососевых в реки, где проводится реинтродукция.
М.И. Базаров, В.К. Голованов
Борок, ИБВВ РАН, 152742, Борок, Ярославской области, Некоузского района,
Смотрите также