
СРАВНЕНИЕ РАННЕГО ОНТОГЕНЕЗА МЕЖДУ ЛОСОСЯМИ АТЛАНТИЧЕСКИХ (РОД SALMO) И ТИХООКЕАНСКИХ (РОД ONCORHYNCHUS) ВИДОВ
Тщательное сопоставление стадий развития эмбрионально-личиночного периода между видами атлантических (род Salmo) и тихоокеанских (род Oncorhynchus) лососей показывает удивительную степень сходства в их осуществлении этого периода онтогенеза. Некоторые особенности в косвенном для зародышей процессе эпиболии (обрастания желтка бластодермой) и в развитии таких второстепенных органов как плавники не приводят, по-видимому, к дивергенции по этим признакам. Создается впечатление, что консерватизм в реализации раннего онтогенеза разных видов и родов лежит в основе анатомо-морфологического консерватизма их взрослых форм, что проявляется в отсутствии надежных морфологических критериев для систематики лососевых рыб. По-видимому, видообразование последних происходит путем развития адаптивных комплексов в процессе освоения новых ареалов, а не за счет морфологической диверсификации. Обсуждается происхождение некоторых важных адаптаций, свойственных лососям рода Oncorhynchus, для иллюстрации подобного пути неморфологического видообразования.
ВВЕДЕНИЕ
Выполненное нами в достаточно давней уже работе тщательное сравнение эмбрионального и личиночного развития разных экологических и таксономических форм из 3-х хорошо обособленных видов лососей рода Salmo (S. salar, S. trutta, S.gairdneri1) обнаружили почти полную тождественность процесса раннего онтогенеза всех этих форм: среди сотен рассмотренных признаков не было выявлено ни одного, который бы отличал их друг от друга (Городилов 1988)2. Такой результат послужил основанием для вывода о тождественности программ развития видов в пределах одного рода. Можно было бы предположить, что морфологические и морфометрические различия внутри- и межвидового уровня возникают уже в постличиночном онтогенезе.
Поскольку в пределах одного рода не удалось выявить каких-либо различий в раннем онтогенетическом развитии разных видов ни по морфологическим признакам, ни по временным соотношениям в формировании признаков, то логически вытекает задача рассмотреть более удаленные по таксономическому статусу объекты. Очевидно, что следующей ступенью должно быть сопоставление на уровне родов, но в пределах одного семейства. Такая возможность появилась в связи с изучением и описанием нами раннего онтогенеза у пяти видов тихоокеанских лососей из рода Oncorhynchus (данные готовятся к пуликации). Сопоставление раннего развития представителей двух родов лососевых рыб Salmo и Oncorhynchus оказалось возможным, потому что все данные были получены по единой методике, сопоставление велось на одних и тех же стадиях и в одинаковом временном масштабе.
Описание стадий развития видов рода Salmo представлено в ряде публикаций (Городилов 1988, 1998; Gorodilov 1996). Точно такие же описания раннего онтогенеза с использованием той же классификации этого периода, разделенного на серии стадий с обозначением каждой серии однотипных стадий подпериодом (в работе 1988 г. подпериоды обозначались еще периодами) с более подробной характеристикой примерно тех же стадий, что и у видов Salmo, были выполнены на пяти видах рода Oncorhynchus (O. keta, O. gorbuscha, O. nerka, O. kisutch, O. masu). Сопоставление этих описаний показало почти такую же стереотипность раннего онтогенеза всех видов этого рода, как это было показано нами раньше при сравнении развития видов рода Salmo. Небольшие внутриродовые различия между видами касались только некоторых провизорных и второстепенных органов.
1 Недавно Международным Номенклатурным Комитетом по представлению американских систематиков (Smith, Stearly 1989) радужная
форель Salmo gairdneri переклассифицирована в состав рода Oncorhynchus под названием O. mikiss.
2 Некоторые отличия были выявлены только в онтогенезе радужной форели S.gairdneri, кстати, подтверждающие ее большую
таксономическую близость к роду Oncorhynchus.
МАТЕРИАЛ И МЕТОДИКА
Икру для исследования тихоокеанских лососей собирали в разных районах Сахалина и Камчатки и всегда доставляли сразу после оплодотворения икры в термоизолированных ящиках с охлаждением обычно за 1-2 суток от места сбора икры до лабораторных инкубаторов в Биологическом институте Санкт-Петербургского государственного университета в Старом Петергофе. Описание одной из моделей инкубатора приведено в нашей работе (Gorodilov 1996). Раньше в этих же инкубаторах были проведены все наблюдения за эмбрионально-личиночным развитием видов рода Salmo.
Сравнение морфологических стадий при развитии разных таксономических групп производили установлением степени сходства ведущего признака, по которому обозначается та или иная стадия, а также комплекса сопутствующих ему признаков. Измерения продолжительности развития выражали в тау-сомитах. Тау-сомит, или τs - время формирования одной пары сомитов в течение равномерного сомитогенеза.
Последний включает в себя у разных видов лососевых образование не менее 60-64 пар сомитов (считая с 1-й пары) с затратой одинакового времени (τs) (Городилов 1988, 1990). После вылупления личинок наблюдали при тех же условиях до полного рассасывания желтка. Относительную длительность стадий в эквивалентах τs рассчитывали как частное от деления τn/τs, где τn — время развития оцениваемых стадий. Относительно других деталей методических условий мы отсылаем к более ранним работам (Городилов 1988; Gorodilov 1996).
РЕЗУЛЬТАТЫ
1. СКОРОСТЬ СОМИТОГЕНЕЗА У РАЗНЫХ ПРЕДСТАВИТЕЛЕЙ
РОДОВ SALMO И ONCHORHYNCHYS
Благодаря очень строгой ритмичности процесса сомитогенеза и его большой протяженности, включающей до 60-64 равновеликих по времени морфогенетических акций, удается определить интервал τs с очень большой точностью. Выяснилось, что зависимость τs от температуры, а в более широком плане и всего эмбриогенеза, хорошо описывается уравнением квадратичной параболы (Городилов 1992). Для всех рассматриваемых видов были определены параметры данной формулы (для видов Salmo см.: Городилов 1992; для видов Oncorhynchus неопубл.). Благодаря этой формуле можно получить значения τs для каждого вида не только в пределах допустимых для зародышей температур, но и за пределами таковых, имеющих только теоретическое значение.
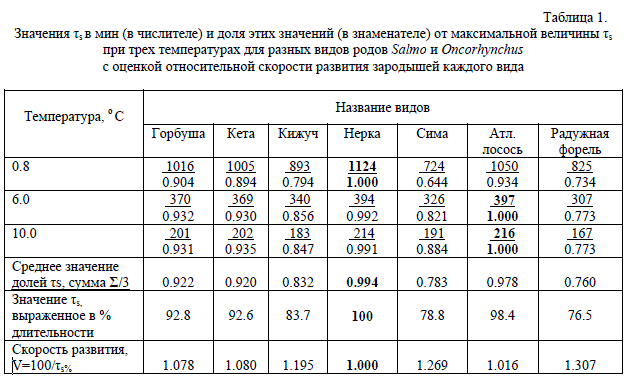
Мы решили сравнить скорости развития разных видов лососей по величине τs. Поскольку наклон графиков параболы для разных видов неодинаковый, то такое сравнение лучше провести для нескольких температур. В табл. 1 приведены данные для трех разных температур: низкой (0.8о С), средней (6.0о С) и высокой (10.0о С). Мы приняли наибольшие значения τs для каждой температуры за 1, рассчитав какую долю по отношению к этому максимуму составляют соответствующие значения τs для каждого вида. Суммировав относительные доли τs для каждой температуры и затем разделив сумму на 3, мы получаем (третья строчка снизу в табл. 1) среднее значение долей τs для всех температур, которое является как бы показателем усредненного значения скорости эмбриогенеза для каждого вида. По этим данным видно, что нерка является видом, у которого наиболее медленно развивающиеся зародыши. Если принять продолжительность τs нерки за 100 % (предпоследняя строка в табл. 1), то усредненная продолжительность этого же интервала у всех остальных видов составляет от 76.5 до 98.4 %. Взяв в расчет, что длительность развития (τs) находится в обратной пропорциональной зависимости со скоростью развития (V), т.е. V= 1/τs•100 %, и принимая скорость развития зародышей нерки за 1, мы можем оценить насколько быстрее нерки развиваются все другие сравниваемые виды. Атлантический лосось имеет более высокий показатель скорости развития, чем нерка, всего на 0.016 (1.6 %), а горбуша и кета на 7.8-8.0 %. Кижуч, сима и радужная форель развиваются быстрее нерки на 20-30 %.
2. ПОДПЕРИОДЫ ДРОБЛЕНИЯ, БЛАСТУЛЫ И ГАСТРУЛЫ
Имеются внутриродовые различия в размерах и в окраске икринок у тихоокеанских лососей, по которым можно приблизительно судить об их видовой принадлежности. Так, икринки у чавычи и кеты наиболее крупные по размерам, а у нерки и симы наиболее ярко окрашены в оранжевый цвет. В этом отношении икринки атлантических лососей рода Salmo не имеют даже такой видоспецифичности. Что же касается процессов развития, то все они, за исключением отмеченных выше различий в скорости развития, протекают совершенно одинаково у всех видов сравниваемых родов (кроме отмеченных раньше различий для радужной форели, см. Городилов 1988). Относительная продолжительность развития от осеменения до начала 124
Atlantic salmon: biology, conservation and restoration, 2003
гаструляции у двух видов рода Salmo (атлантический лосось и кумжа) и у пяти исследованных видов рода Oncorhynchus составляет 34-36 эквивалентов τs, причем общая продолжительность подпериодов оплодотворения и дробления составляет, по-видимому, 12 (Городилов 1988), а продолжительность подпериода бластуляции 22-24 τs. Относительная продолжительность подпериода гаструляции, заканчивающегося перед началом сомитогенеза, одинакова у всех сравниваемых видов и составляет примерно 20±1 τs.
Таблица 1.
Значения τs в мин (в числителе) и доля этих значений (в знаменателе) от максимальной величины τs
при трех температурах для разных видов родов Salmo и Oncorhynchus
с оценкой относительной скорости развития зародышей каждого вида
3. ПОДПЕРИОД СОМИТОГЕНЕЗА
В течение этого подпериода происходит закладка и частичное развитие почти всех основных органов и функциональных систем. Начало закладок, как и некоторые моменты их последующего морфогенеза, удобно соотнести с числом сомитов, и наличие этого показателя позволяет сопоставить эти ключевые моменты в развитии разных органов между разными видами. Выяснилось, что у зародышей всех видов обоих сравниваемых родов появление закладок одинаковых органов или сходные морфологические преобразования всегда сочетаются с одним и тем же числом сомитов. Об этом списке признаков, появляющихся в течение сомитогенеза, можно справиться в более ранних наших работах (Городилов 1988, 1998). Каких-либо морфологических особенностей или гетерохроний развития представленного набора признаков установить не удалось: развитие всех видов происходит как бы по одной и той же программе, настолько постоянными оказываются не только комплексы признаков, но и последовательность их развития. Сходным оказывается и общий возраст зародышей, выраженный в τs, для стадий с одинаковым количеством сомитов. Однако имеются две особенности в развитии зародышей в течение этого подпериода, по которым можно различить хотя бы часть видов рода Oncorhynchus от видов рода Salmo.
Первой особенностью здесь оказались данные, связанные с процессом эпиболии или обрастания желточного мешка слоем бластодермы. Этот процесс начинается в конце бластуляции-начале гаструляции после того как образуется зародышевое кольцо по краю бластодиска, которое постепенно распространяется от анимального полюса к вегетативному и тянет за собой клеточный слой бластодермы. Поскольку у лососевых процесс эпиболии обычно совпадает с сомитогенезом зародышей, то разные стадии эпиболии удобнее всего идентифицировать по числу пар сомитов. В табл. 2 мы приводим данные о стадиях развития собственно зародыша, выраженным числом сомитов, соотнесенным со стадией завершения процесса эпиболии у всех видов Oncorhynchus и некоторых представителей рода Salmo.
Таблица 2.
Количество сомитов у зародышей разных видов и популяций лососевых рыб
из родов Salmo и Oncorhynchus к стадии завершения процесса эпиболии
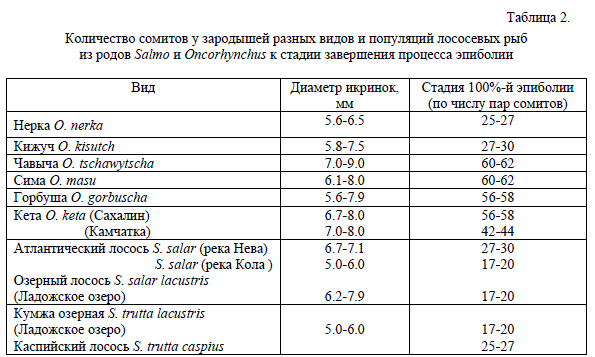 Хотя некоторыми авторами отмечается, что стадия, на которой завершается процесс эпиболии, у зародышей из одной популяции или даже от одной самки зависит от диаметра желточного мешка (Коровина 1978; Игнатьева 1979; Городилов 1982), но обычное варьирование в пределах одной популяции достигает здесь величины всего нескольких сомитов. В то же время из данных таблицы ясно, что виды из рода Oncorhynchus по этому признаку разделяются на две четко обособленные группы: в одной находятся кижуч и нерка, у которых эпиболия завершается на стадиях 25-30 пар сомитов, а в другой чавыча, сима, горбуша и сахалинская кета, у которых окончание эпиболии приходится на стадии 56-62 пар. С другой стороны, различия наблюдаются и на внутривидовом уровне между сахалинской и камчатской кетой, а также внутри вида Salmo salar между балтийской и кольской популяциями, где различия в стадиях зародыша в конце эпиболиии достигают 10-15 пар сомитов.
Хотя некоторыми авторами отмечается, что стадия, на которой завершается процесс эпиболии, у зародышей из одной популяции или даже от одной самки зависит от диаметра желточного мешка (Коровина 1978; Игнатьева 1979; Городилов 1982), но обычное варьирование в пределах одной популяции достигает здесь величины всего нескольких сомитов. В то же время из данных таблицы ясно, что виды из рода Oncorhynchus по этому признаку разделяются на две четко обособленные группы: в одной находятся кижуч и нерка, у которых эпиболия завершается на стадиях 25-30 пар сомитов, а в другой чавыча, сима, горбуша и сахалинская кета, у которых окончание эпиболии приходится на стадии 56-62 пар. С другой стороны, различия наблюдаются и на внутривидовом уровне между сахалинской и камчатской кетой, а также внутри вида Salmo salar между балтийской и кольской популяциями, где различия в стадиях зародыша в конце эпиболиии достигают 10-15 пар сомитов.Другим существенным признаком, который отличает уже все виды рода Oncorhynchus от видов рода Salmo, являются максимальные числа пар сомитов, которые образуются в течение сомитогенеза у тех и других видов. В табл. 3 можно видеть, что у атлантических лососей (т.е. у S. salar и S. trutta) максимальное число сомитов 65-68 и никогда их не образуется 70 или больше.
Таблица 3.
Максимальное число пар сомитов, образующихся в процессе сомитогенеза
у разных видов рыб из родов Salmo и Oncorhynchus
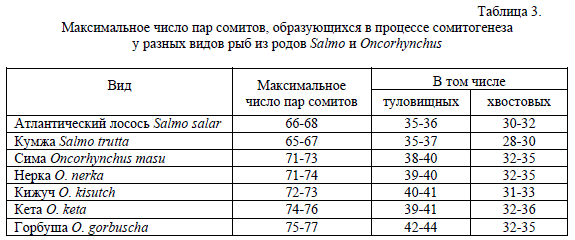
В то же время у всех тихоокеанских лососей сомитогенез завершается образованием не менее 70 пар сомитов и в результате у них образуется этих структурных единиц минимум на 3-6 и максимум на 7-9 больше, чем у атлантических. Интересно, что у тихоокеанских лососей анус, относительно положения которого сомиты делятся на туловищные и хвостовые, сдвинут более каудально по сравнению с его положением у атлантических лососей, особенно у горбуши, хотя пропорции тела, по-видимому, сохраняются за счет увеличения числа хвостовых сомитов. Заметим, что часть хвостовых сомитов затем редуцируется и конечное число метамеров (сегментов, позвонков и пр.) меньше числа образуемых сомитов.
4. ПОДПЕРИОД ВАСКУЛЯРИЗАЦИИ ЖЕЛТОЧНОГО МЕШКА
В конце периода сомитогенеза происходит образование желточной вены, которая затем формирует на поверхности желточного мешка мощную капиллярную сеть. Наряду с этим в глазных бокалах начинается синтез меланина, гранулы которого постепенно появляются сначала по краям бокалов, а затем распространяются по их сфере. Приведем характеристику основных признаков, отмеченных в период васкуляризации у лососей двух сравниваемых родов (табл. 4).
Весь подпериод васкуляризации желтка длится около 60 τs. В табл. 4 мы приводим описание лишь части внешних признаков, которые развиваются сходным образом у всех видов, в то же время стремясь указать по возможности все различия, которые нам удалось выявить. В основном эти различия касаются второстепенных признаков и процессов, которые не играют существенной роли для развития зародыша, таких, например, как различия в скорости освобождения головы от желтка. Более существенной для онтогенеза можно считать задержку закладки плавников Д и А, поскольку эти плавники у лососей из рода Salmo начинают формироваться в этом подпериоде, тогда как у видов рода Oncorhynchus этот момент сдвинут на следующий подпериод.
5. ПОДПЕРИОД ОБРАЗОВАНИЯ ЛУЧЕЙ-ЛЕПИДОТРИХИЙ В ХВОСТОВОМ ПЛАВНИКЕ Вскоре после образования скопления мезенхимных клеток в хвостовом плавнике, здесь начинают
формироваться лучи-лепидотрихии, которые закладываются по одному с интервалом примерно равным 7 τ
Веер из 21-22 лучей образуется к концу подпериода. Весь этот подпериод является достаточно протяженным,
продолжаясь около 150-160 τ Его начало у тихоокеанских лососей можно идентифицировать по закладке
сначала анального, а затем дорзального плавников, в то время как у атлантических лососей закладка этих же
плавников происходит одновременно и раньше на 10-15 τ В табл. 5 мы приводим данные по возрасту
закладки не только этих двух непарных плавников, но и обоих парных, для того, чтобы наглядно сопоставить
сходства и различия в развитии этих органов у всех сравниваемых видов
s. s. s.
Из табл. 5 мы видим, что закладки всех 4-х типов плавников у обоих видов рода Salmo происходят на одних и тех же стадиях. У всех видов рода Oncorhynchus оба парных плавника закладываются на той же или практически на той же стадии. Последующее после закладки А, Д и брюшных плавников развитие зародышей у тихоокеанских и атлантических лососей в основном совпадает. Расхождения начинаются перед началом вылупления и они не связаны с морфологическими структурами, а скорее с различиями в функциональном состоянии зародышей двух родов. Эти различия проявляются в различном отношении атлантических и тихоокеанских лососей к температуре инкубации перед вылуплением и в процессе вылупления зародышей из оболочек. Оказалось, что на поздних стадиях эмбриогенеза зародыши тихоокеанских лососей способны замедлять развитие при высоких температурах. Это явление было обнаружено нами при измерениях возраста зародышей в относительных единицах во время периода вылупления. Достаточно известно, что начало вылупления зародышей не приурочено к определенной морфологической стадии и может сдвигаться на другие стадии в зависимости от разных факторов и особенно от температуры. У атлантических лососей, как и у многих других рыб, повышение температуры сдвигает вылупление зародышей на более ранние стадии, т.е. стимулирует этот процесс (табл. 6).
Оказалось, что у тихоокеанских лососей высокая температура не только не стимулирует процесс вылупления зародышей, но даже замедляет его начало, причем в тем большей степени, чем выше температура. Наиболее ярко это замедление проявляется у горбуши. Приведем табл. 7 относительной продолжительности развития горбуши от оплодотворения до начала, середины и конца периода вылупления.
Очевидно, что чем выше температура в инкубаторе, тем в более позднем возрасте происходит вылупление: минимальный возраст массового вылупления у горбуши 330 τs (1о), максимальный 600 τs (14.4о).
Таблица 4.
Возраст и характеристика стадий развития зародышей рыб из родов Salmo и Oncorhynchus
в подпериоде васкуляризации желточного мешка
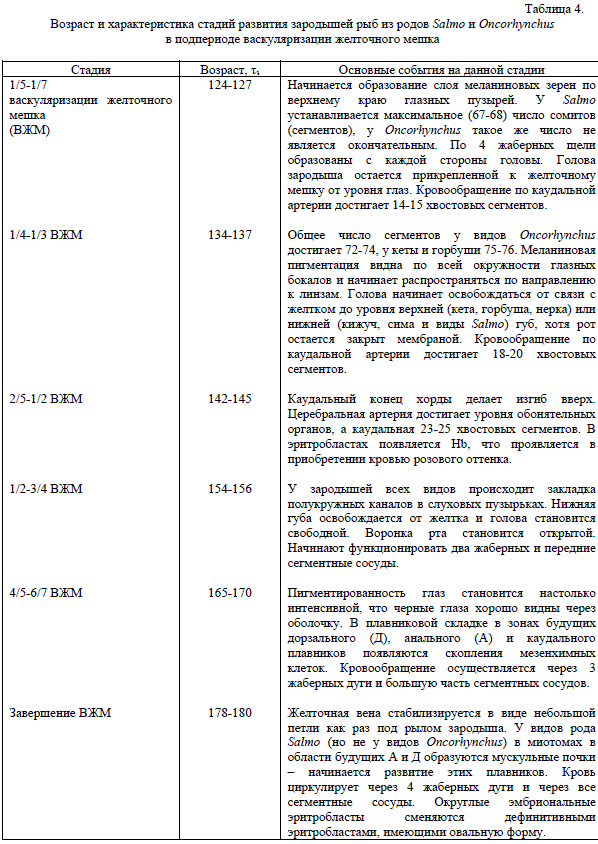 Начинается образование слоя меланиновых зерен по верхнему краю глазных пузырей. У Salmo устанавливается максимальное (67-68) число сомитов (сегментов), у Oncorhynchus такое же число не является окончательным. По 4 жаберных щели образованы с каждой стороны головы. Голова зародыша остается прикрепленной к желточному мешку от уровня глаз. Кровообращение по каудальной артерии достигает 14-15 хвостовых сегментов.
Начинается образование слоя меланиновых зерен по верхнему краю глазных пузырей. У Salmo устанавливается максимальное (67-68) число сомитов (сегментов), у Oncorhynchus такое же число не является окончательным. По 4 жаберных щели образованы с каждой стороны головы. Голова зародыша остается прикрепленной к желточному мешку от уровня глаз. Кровообращение по каудальной артерии достигает 14-15 хвостовых сегментов.
Общее число сегментов у видов Oncorhynchus достигает 72-74, у кеты и горбуши 75-76. Меланиновая пигментация видна по всей окружности глазных бокалов и начинает распространяться по направлению к линзам. Голова начинает освобождаться от связи с желтком до уровня верхней (кета, горбуша, нерка) или нижней (кижуч, сима и виды Salmo) губ, хотя рот остается закрыт мембраной. Кровообращение по каудальной артерии достигает 18-20 хвостовых сегментов.
Каудальный конец хорды делает изгиб вверх. Церебральная артерия достигает уровня обонятельных органов, а каудальная 23-25 хвостовых сегментов. В эритробластах появляется Hb, что проявляется в приобретении кровью розового оттенка.
У зародышей всех видов происходит закладка полукружных каналов в слуховых пузырьках. Нижняя губа освобождается от желтка и голова становится свободной. Воронка рта становится открытой. Начинают функционировать два жаберных и передние сегментные сосуды.
Желточная вена стабилизируется в виде небольшой петли как раз под рылом зародыша. У видов рода Salmo (но не у видов Oncorhynchus) в миотомах в области будущих А и Д образуются мускульные почки – начинается развитие этих плавников. Кровь циркулирует через 4 жаберных дуги и через все сегментные сосуды. Округлые эмбриональные эритробласты сменяются дефинитивными эритробластами, имеющими овальную форму.
128
Atlantic salmon: biology, conservation and restoration, 2003
Таблица 5.
Относительная продолжительность развития различных типов плавников
у разных видов из родов Salmo и Oncorhynchus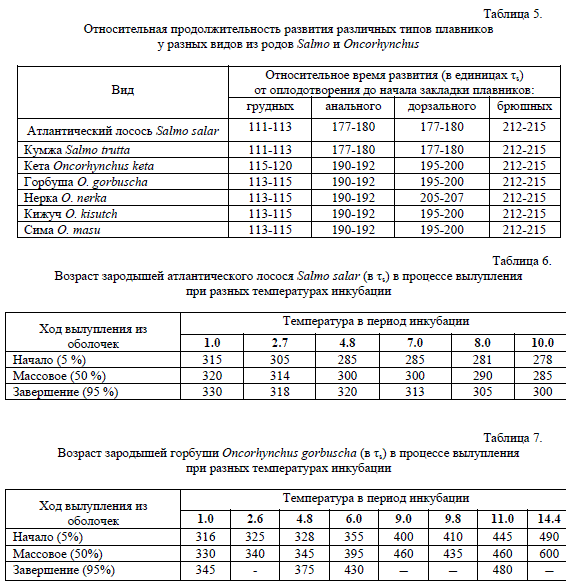
6. ПОДПЕРИОД ОФОРМЛЕНИЯ НЕПАРНЫХ И ПАРНЫХ ПЛАВНИКОВ
В этот подпериод мы включили все развитие после образования полного веера из лучей-лепидотрихий в хвостовом плавнике (340-360 τs) до окончательного рассасывания желточного мешка. У атлантического лосося раньше мы выделяли подпериод свободных эмбрионов (от 340 до 450 τs) и личиночный период развития (от 450 до 600 τs) (Городилов 1998). Теперь мы решили упростить классификацию этой части онтогенеза, выделив процесс оформления плавников как один из подпериодов в составе всего эмбрионально-личиночного периода.
В этом подпериоде наиболее заметные морфологические процессы связаны с преобразованиями плавниковой складки вокруг тела и выделением из ее состава Д, А а также жирового (Ж) плавника. Все эти процессы происходят достаточно одинаково и в сходной последовательности. Различие между видами Salmo и Oncorhynchus заключается в основном лишь в том, что у первых они проходят в более раннем возрасте, чем у вторых. Различия также наблюдаются в процессах пигментации покровов тела и заключаются они в том, что в разном возрасте и в разных частях тела могут преобладать пестряточная (меланофоры), серебристая (иридофоры) или золотистая (ксантофиллы) окраски.
Таблица 8.
Возраст и характеристика стадий развития зародышей и личинок рыб из родов Salmo и Oncorhynchus
в подпериоде оформления непарных и парных плавников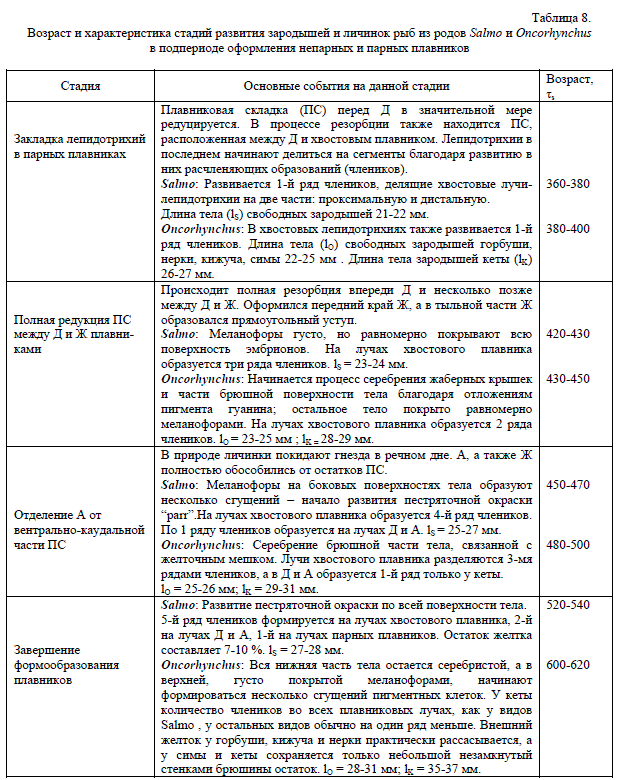
ОБСУЖДЕНИЕ
В данном исследовании произведено детальное сравнение раннего развития рыб семейства лососевых, принадлежащих к двум родам Salmo и Oncorhynchus. Предварительное исследование подобного плана было произведено только на видах рода Salmo, когда один из авторов (Городилов 1988) сравнивал эмбриональное и личиночное развитие разных экологических и таксономических форм атлантического лосося S. salar, кумжи 130
Atlantic salmon: biology, conservation and restoration, 2003
S. trutta, а также относимой в тот период к этому же роду радужной форели S. gairdneri. В развитии первых двух видов была обнаружена полная тождественность в процессах раннего онтогенеза, причем ни одного признака, который бы их отличал друг от друга в этот период онтогенеза, не было обнаружено. Что касается радужной форели, то в ее развитии был выявлен единственный существенный признак таксономического ранга, который отличал ее от других видов в этом роде, — общее число сомитов, формировавшихся в процессе сомитогенеза, у зародышей радужной форели было на 3-6 пар больше, чем у кумжи и атлантического лосося. Этот признак сближал радужную форель больше с тихоокеанскими Oncorhynchus, чем с атлантическими Salmo (Городилов 1988). С учетом всего этого представляло интерес сравнить периоды ранних онтогенезов у разных видов из двух этих родов. Наши исследования по описанию раннего онтогенеза у пяти видов лососей из рода Oncorhynchus предоставили такую возможность.
В результате тщательного сопоставления стадии за стадией всего эмбрионального и личиночного развития двух видов из рода Salmo и пяти видов из рода Oncorhynchus мы приходим к выводу о необычайно высоком уровне сходства как по отдельным признакам и комплексам признаков для каждой из стадий, так и по пропорциям времени, необходимого для образования большинства сопоставляемых признаков. Этот вывод основывается на описании и сравнении не менее 40 стадий, не считая значительного количества промежуточных стадий, описание которых специально не производили, но прослеживали в процессе развития. Можно говорить о типажности серий стадий развития и целых комплексов морфологических и функциональных признаков на этих стадиях, об их стабильности в хронологической очередности и в относительных пропорциях между собой, и это наблюдается почти на всем протяжении эмбрионально-личиночного развития. В то же время некоторые различия между этими родами были выявлены.
В первую очередь здесь нужно отметить различия между родами по максимальному числу пар сомитов, которые образуются в результате сомитогенеза. Эти данные сведены в табл. 3. Здесь можно видеть, что у зародышей атлантических лососей количество этих метамеров образуется меньше, чем у разных видов тихоокеанских лососей на величину от 3-6 до 7-9. Именно этот признак сближает радужную форель с лососями из рода Oncorhynchus и данный факт может стать еще одним аргументом в пользу включения форели в этот род. Другой признак, связанный с разным числом сомитов у тех и других видов, касается увеличения числа туловищных сегментов за счет сдвига положения ануса у зародышей Oncorhynchus в каудальную часть тела в основном на 3-5, а у горбуши даже на 6-8 сегментов. Интересно, что у тихоокеанских видов увеличивается также число хвостовых сегментов, но этот признак не является стабильным, поскольку несколько наиболее каудальных сегментов позднее деградируют.
Довольно интересными оказались также данные, связанные с процессом эпиболии или обрастания желточного мешка слоем бластодермы. Сам процесс обрастания в значительной мере протекает автономно и, по-видимому, не скоррелирован с процессами формирования собственно зародыша. Продолжительность и окончание процесса эпиболии зависят от размеров желточного мешка и не являются однозначными у особей даже из одной популяции, но варьирование здесь обычно не превышает 3-5 сомитов. В то же время даже в пределах вида стадия, на которой может находиться собственно зародыш на момент окончания процесса эпиболии, оказывается настолько различающейся, что объяснение этих расхождений только размерами икринок было бы явным упрощением. Различия между сахалинской и камчатской популяциями кеты, а также внутри вида Salmo salar между балтийской и кольской популяциями в конце эпиболии достигают 10-15 пар сомитов (табл. 2). Еще большие различия по этому признаку достигаются между разными видами рода Oncorhynchus. В этом роде четко выделились две группы видов. В одной группе находятся кижуч и нерка, у которых эпиболия завершается на стадиях 25-30 пар сомитов, а в другой — чавыча, сима, горбуша и сахалинская кета, у которых этот процесс завершается на стадиях 56-62 пар.
Некоторые авторы использовали различия в стадиях зародышей родственных видов, на которых заканчивался процесс эпиболии, для построения спекулятивных филогенетических схем (Коровина 1978; Решетников 1980). Было бы заманчиво на основании полученных фактов, связанных со скоростью эпиболии, попытаться выстроить также некие филогенетические связи. Однако накопленный нами материал по многим видам костистых рыб свидетельствует о малой ценности таких построений. Поэтому мы предпочтем воздержаться от теоретизирования, предполагая, что процесс эпиболии настолько автономен от развития собственно зародыша, что он может легко изменять свою скорость и завершаться быстрее или медленнее, до определенного времени, не влияя на развитие последнего. Видимо бластодермальный слой начинает играть свою более важную роль с началом процесса развития желточной кровеносной системы, т.е. васкуляризации желточного мешка. В то же время было бы интересно в будущем выяснить, почему две группы тихоокеанских лососей очень резко отделились друг от друга по рассматриваемому признаку.
При определении стадий закладки разных типов плавников нам удалось выявить довольно четкие гетерохронии по возрасту закладки анального и дорзального непарных плавников у тихоокеанских лососей по сравнению с атлантическими (табл. 5).
Согласно наиболее популярным в современном эволюционизме взглядам считается, что основной причиной филогенетических преобразований являются возникающие в онтогенезе гетерохронии во времени закладки, в скорости развития и т.д., тех или иных признаков (Gould 1977; Gilbert et al. 1996). Предполагается,
131
Atlantic salmon: biology, conservation and restoration, 2003
что небольшие изменения в раннем онтогенезе могут, каскадно увеличиваясь, вызвать большие фенотипические перестройки в дефинитивных признаках (Gould 1977, 1992). Выявив столь редкие, как показали данное и предыдущее (Городилов 1988) наши исследования, в онтогенезе лососевых гетерохронии, можно было бы ожидать каких-либо перестроек или изменений в плавниках у представителей сравниваемых родов. Однако известно, что систематики никогда не выделяют каких-либо особенностей в строении или форме плавников у этих лососей, и, следовательно, описанные гетерохронии не влияют на их дефинитивное состояние.
Процесс расхождения в направленности и в уровне дифференцировки между видами Salmo и Oncorhynchus в какой-то мере начинается после вылупления зародышей из оболочки яйца в подпериоде формирования плавников (табл. 8). Это проявляется в некотором отставании развития членистой структуры опорных лучей-лепидотрихий во всех типах плавников у одноразмерных личинок лососей из рода Oncorhynchus по сравнению с личинками рода Salmo. Другое отличие связано с тем, что у личинок лососей из рода Salmo начинает развиваться пестряточная окраска, тогда как у тихоокеанских лососей проявляется признак ранней смолтификации — развитие “серебристой” окраски тела. Данное отличие связано с обязательностью или возможностью ската тихоокеанских лососей в море уже в личиночный период.
Итак, детальное сопоставление эмбрионального и личиночного развития лососей из двух близкородственных родов обнаружило высокий уровень сходства, а все выявленные различия связаны либо с провизорными органами, либо не учитываются в таксономических работах (как, например, число туловищных сегментов). Можно было бы предположить, что реальные морфологические и морфометрические различия внутри- и межвидового уровня возникают в постличиночном онтогенезе. Однако знакомство с работами, посвященными оценкам систематического статуса рыб семейства лососевых, показывает, что среди географических, сезонных. кариотипических критериев видовой обособленности наименее четкими и наиболее спорными являются анатомо-морфологические признаки. Например, для различения разных видов рода Salmo и подрода Parasalmo, который раньше входил в данный род, после долгих исследований удалось выделить 4 остеологических признака в хондрокраниуме. Это касается некоторых деталей строения сошника, Supraethmoideum и еще двух костей (Дорофеева 1985). Знакомство с представленными иллюстрациями образцов (с. 6)оставляет впечатление малости различий в форме этих костей, хотя мы не пытаемся ставить под сомнение их наличие. Мы просто хотим подчеркнуть удивительный уровень консервативности, который проявляется в сохранении даже деталей строения хондрокраниума и всего скелета в пределах не только родов, но и, видимо, более крупных таксонов. Систематики, которые всегда занимаются поисками различий, это свойство оставляли обычно без внимания. Нам же остается связать между собой две серии фактов: консервативность анатомического строения взрослых рыб из разных видов и родов лососевых логически вытекает из консервативности индивидуального развития всех этих видов. Иначе говоря, мы не находим существенных отличий в морфологии взрослых рыб, потому что они проистекают из сходства их индивидуального развития.
Проблема таксономии лососевых рыб активно разрабатывается свыше 100 лет и всегда представляла
трудную задачу, поскольку многие виды имеют более или менее широкую перекрываемость всех
пластических и меристических признаков. Это отсутствие четких разделительных границ можно объяснить
следующим образом.
Уже ранние предковые виды лососевых выработали достаточно совершенную форму тела, что позволяет
им находиться до сих пор в состоянии биологического прогресса (Гербильский 1967). Дальнейшее
совершенствование и эволюция видов происходят на пути выработки новых адаптаций и затрагивают в
большей мере не анатомо-морфологические, а функциональные системы организма. Соответственно,
таксономическая обособленность форм может осуществляться не на уровне анатомо-морфологических
признаков, а на уровне комплексов адаптаций, возникающих в изолированных популяциях в конкретной для
них эколого-географической среде обитания.
Процесс такого видообразования можно представить в виде ряда последовательных этапов:
1) Выделение в пределах исходного вида двух или нескольких изолированных географически популяций, т.е. получение аллопатрического статуса, который является обязательным условием видообразования (Майр 1968).
2) Освоение новой эколого-географической зоны, ее пищевых ресурсов, приспособление к климатическим условиям, перестройка стратегии жизненного цикла.
3) По ходу этого освоения происходит выработка новых комплексов адаптаций, которые совершенствуют условия жизни в новой реальной среде.
4) Одновременно генетико-автоматические процессы могли обеспечить перестройку кариотипов и закрепить изоляцию вида между бывшими популяциями одного вида уже на цитологическом уровне.
5) В тех случаях, когда генетические перестройки достигнут такого уровня, что способны будут обеспечить генетическую изоляцию между двумя такими новыми видами, они могут вновь совместиться в одной и той же эколого-географической среде, т.е. опять стать симпатричными.
132
Atlantic salmon: biology, conservation and restoration, 2003
6) Сосуществование видов в одном ареале поддерживает и дальше отбор в сторону дальнейшего усиления генетической репродуктивной изоляции
Последняя ситуация хорошо прослеживается на некоторых парах видов, которые являются близкими по занимаемому ими ареалу, по жизненному циклу, спектру питания, местам нереста и т.д. Такой парой определенно являются атлантический лосось и кумжа. Эти оба вида имеют огромные ареалы, значительная часть которых совпадает. Они сосуществуют в одних и тех же биоценозах, по сути, являясь пищевыми конкурентами и конкурентами на нерестилищах. В то же время известно, что по кариотипам они имеют самые существенные среди лососевых различия. S. trutta имеет 100-102 хромосомных плеча (NF) в диплоидном наборе. S.salar занимает совершенно особое место во всем подсемействе, имея NF равным всего 72-74. Соответственно модальное число хромосом у кумжи 2n = 80, причем из них 20 мета- и субметацентрических (m/sm), или двуплечих, и 60 тело- и субтелоцентрических (t/st), или одноплечих; у атлантического лосося число хромосом 2n =54-58, из них 14-18 m/sm и 38-46 t/st (Викторовский 1978; Hartley & Horne 1984а, b; Phillips, Rab 2001).
Другой подобной парой являются тихоокеанские лососи — кета и горбуша. Из 6 видов рода Oncorhynchus эти виды действительно наиболее близко сосуществуют и особенно ожесточенно они конкурируют на нерестилищах. Кроме того, эти виды наиболее жестко специализированы по своему жизненному циклу, что проявляется в раннем, еще в личиночном возрасте, скате в море и в отсутствии жилых форм. В то же время эти виды имеют наибольшие среди тихоокеанских лососей отличия в структуре кариотипов. Несмотря на то, что оба вида, как и все остальные среди рода Oncorhynchus, имеют в кариотипе практически одинаковое число NF (100-104), структура их хромосом является весьма различной. Показано, что у горбуши число хромосом равно 52 и все они являются метацентриками (Горшков, Горшкова 1981; Phillips, Ihssen 1985). У кеты число хромосом 2n = 74, из которых 26 m и 48 t/st (Phillips, Ihssen 1985; Phillips, Rab 2001). Остальные 4 вида по соотношению мета- и телоцентрических хромосом занимают промежуточное положение между кетой и горбушей. Согласно Р. М. Викторовскому (1978), эволюция кариотипов в роде Oncorhynchus шла в основном по пути центрических слияний и у наиболее продвинутой и крайне специализированной горбуши все возможные преобразования кариотипа путем центрических слияний уже исчерпаны, т.к. все хромосомы стали метацентриками. Стоящая рядом с горбушей по уровню специализации кета, в то же время стоит на другом полюсе этой группы видов по структуре кариотипов: кета имеет наименьшее число метацентриков.
Существует мнение, что у лососевых между количеством хромосом (при значительной стабильности чисел NF) и разнообразием стратегий жизненного цикла существует прямая связь: виды, имеющие большие числа 2n (т.е. в их хромосомном наборе имеется много t/st) имеют более обширный ареал и больше вариантов жизненной стратегии (озерные, речные, анадромные), чем виды с меньшими 2n (в их хромосомных наборах больше всего хромосом типа m). Эти последние считаются обычно жестко специализированными анадромными видами (Викторовский 1978; Phillips, Rab 2001). Связаны ли между собой эволюция морфофизиологических адаптаций и эволюция кариотипов или они изменяются независимо друг от друга? В принципе независимость эволюции генетического материала может базироваться на открытых еще в 1930-х годах Н. П. Дубининым и С. Райтом (S. Wright) генетико-автоматических процессах. Однако вряд ли существует полная независимость эволюции кариотипов от факторов естественного отбора. Таковым фактором, поддерживаемым естественным отбором, может быть создание генетической изоляции в аллопатрических популяциях вначале на границах их ареалов, а затем по мере совмещения мест обитания, ее последующее усиление. Так, видимо, происходило в случае рассматриваемых нами видов, у которых отбор определил развитие генетико-автоматических процессов в противоположных направлениях, а именно: у горбуши в сторону слияния телоцентрических хромосом и увеличения числа метацентриков, а у кеты в сторону разделения метацентриков и уменьшения их числа.
Тот факт, что у этих наиболее близко сосуществующих видов имеются наибольшие различия в структуре кариотипов, что обеспечивает наибольшую цитологическую изолированность, позволяет сформулировать следующее правило: чем менее географически изолированы родственные виды, тем больше они должны быть изолированы генетически за счет перестройки структуры кариотипов. Или, иначе говоря: Условием, согласно которому два географически изолированных вида могут начать вновь сосуществовать в единой географической зоне, должна быть достаточно надежная цитологическая или генетическая несовместимость.
Рассмотрим некоторые комплексы адаптаций, которые отличают тихоокеанских лососей от атлантических и попытаемся выяснить причины, которые привели к их возникновению в конкретной эколого-географической обстановке региона Пацифики. Как считают, род Oncorhynchus произошел от тихоокеанских лососей подрода Parasalmo, которые обитают в наиболее северных районах Пацифики (Neave 1958; Vladykov 1963). Завоевание новых ареалов происходило по пути продвижения на юг. Популяции Parasalmo по мере распространения на юг нашли здесь чрезвычайно благоприятные условия для увеличения численности. Это и мягкий климатический режим и чрезвычайно благоприятные условия для нагулов в океане с изобилием корма. Эти условия, а также свойственная лососевым забота о потомстве, обеспечивали постоянное повышение численности популяций, которая стала лимитироваться ограниченным количеством нерестовых 133
Atlantic salmon: biology, conservation and restoration, 2003
площадей и низкой кормностью существующих здесь пресноводных водоемов. Предки онкоринхусов освоили постепенно все реки, речки и ручейки по азиатскому и американскому побережьям Тихого океана. В то же время неисчерпаемые запасы кормовых ресурсов в океане все более увеличивали численность нерестовых стад, возвращающихся в реки. Чтобы разрешить это противоречие, на этом пути стали вырабатываться механизмы посленерестовой деградации и гибели нерестующих рыб. Рыбы после нереста в большинстве гибли в любом случае, поскольку не могли найти корма в реке. Впоследствии эти механизмы возникли уже как обязательные элементы жизненного цикла.
С другой стороны, огромное количество отложенной икры создавало проблему, когда весной появлялась
масса личинок, которые были не в состоянии прокормиться в реке. Только немедленный скат в море, где
имеется богатая кормовая база, мог обеспечить шансы для выживания. Это стимулировало развитие
механизмов личиночной смолтификации. Такая реконструкция развития данной адаптации подтверждается
тем, что у наиболее многочисленных видов — горбуши и кеты — этот механизм, обеспечивающий скат
личинок в море, является почти облигатным. Такие же виды как нерка, чавыча, кижуч, сима имеют меньшую
численность, и у них механизм личиночного ската в море регулируется в зависимости от численности
популяции. Например, более многочисленная чавыча американского побережья мигрирует в океан уже в
первое лето жизни, тогда как на азиатском побережье она малочисленна и остается в пресной воде не менее
года (Леванидов 1976).
Еще одной необычной адаптацией, возникшей у тихоокеанских лососей и отсутствующей у атлантических
лососей и у многих других рыб, является способность зародышей из рода Oncorhynchus замедлять развитие
при высоких температурах. Явление замедления относительных темпов развития при высокой температуре
можно рассматривать как определенное биологическое приспособление к сезонным циклам природы,
сложившееся в связи с особенностями обитания и размножения этих рыб. Как известно, рыбы видов рода
Oncorhynchus в основной массе нерестятся в конце лета - в первой половине осени, когда температура воды в
реках может быть на уровне 15-18 Смирнов 1975). При такой температуре развитие до начала вылупления
зародышей, скажем, у горбуши, нерестящейся в конце августа, занимало бы 30-35 суток с затратой почти
половины запасов желтка. Если бы не было механизма замедления относительной скорости развития, то
личинкам горбуши просто не хватило бы запасов желтка, чтобы дожить до весны. Возможно, что путем
замедления или снижения метаболизма, зародыши впадают в состояние, в чем-то напоминающее
анабиотическое, хотя в отличие от обычного анабиоза оно вызывается не понижением температуры, а ее
повышением. Благодаря этому развитие не форсируется при тепловых сезонных аномалиях и запасов желтка
хватает на весь зимне-весенний период.
ВЫВОДЫ
1. Сравнение эмбрионально-личиночного развития у разных видов лососевых из родов Salmo (3 вида) и Oncorhynchus (5 видов) обнаружило практически полную тождественность в реализации программы раннего онтогенеза у всех видов обоих родов.
2. Небольшие различия, которые были обнаружены, связаны либо с косвенным для развития самого зародыша процессом эпиболии или сдвигом во времени закладок таких второстепенных органов как плавники. По-видимому, эти особенности онтогенеза не трансформируются в какие-либо дивергентные изменения таксономического ранга.
3. Консерватизм в осуществлении онтогенетических программ на видовом и родовом уровнях, по-видимому, лежит в основе анатомо-морфологического консерватизма, характерного для взрослых организмов у Salmoninae; как известно это подсемейство характеризуется отсутствием надежных морфологических критериев для таксономической идентификации.
4. Отсутствие заметной морфологической изменчивости у четко обособленных видов салмонид могло бы означать, что видообразование у этих видов происходило через развитие комплексов специфических адаптаций, которые существенно преобразовали жизненный цикл.
5. Делается попытка реконструкции в развитии наиболее важных адаптаций у тихоокеанских лососей.
Работа поддержана грантом Минобразования в области естественных наук, рег. №075-0602-281-21.
Ю.Н. Городилов, Е.Л. Мельникова
Биологический институт Санкт-Петербургского государственного университета
Смотрите также