О принципах формирования горбушей анадромных миграционных потоков
По объему добычи среди тихоокеанских лососей естественного воспроизводства горбуша занимает ведущее место. Особенно заметно ее преобладание в уловах из азиатской части ареала (Шунтов, 1994а).
Основные нагульные скопления азиатская горбуша образует в при-курильских океанических водах (преимущественно стада бассейна Охотского моря) и приалеутских океанических водах (преимущественно стада бассейна Берингова моря) (Takagi е! а1., 1981; Ogura, 1994). Отсюда она совершает анадромные нерестовые миграции к побережьям, где в основном и ведется ее промысел.
Научной основой рационального использования запасов рыб является прежде всего знание популяционной структуры вида и его численности.
К настоящему времени в отношении популяционной организации горбуши утвердились следующие представления. На первом структурном уровне выделяются поколения четных и нечетных лет, имеющие устойчивые генетические различия. Это определяется тем, что подавляющее большинство особей созревает на втором году жизни (3-летки и годовики среди производителей отмечены лишь в единичных случаях). Таким образом, существуя в пределах единого ареала, эти поколения оказываются практически изолированными друг от друга. Численность особей четных и нечетных поколений у одного стада может быть близкой, у другого - резко различаться. Такое преобладание способно сохраняться десятилетиями, прежде чем сменится доминанта.
Из всех объяснений возникновения доминирующих поколений правдоподобными признаны два. И.Б.Бирман (1985) предположил воздействие квазидвухлетнего цикла солнечной активности как климатообра-зующего фактора, определяющего различия условий воспроизводства. Л.А.Животовский с соавторами (1996) высказали сомнение в том, что условия воспроизводства в четные и нечетные годы проявляются существенно по-разному. Они попытались отыскать причину доминирования поколений и смены доминант во "внутренних" свойствах системы, моделируя динамику численности горбуши. Их эксперименты в целом оказались удачными. Однако известно, что процессы, протекающие в реальной системе и ее упрощенных подобиях, далеко не всегда адекватны.
На втором структурном уровне (внутри поколений четных или нечетных лет) горбуша имеет уже другие характерные особенности попу-ляционной организации: неустойчивость различий стад по генным маркерам, нечеткость границ между стадами по комплексам морфологических признаков, отсутствие региональной приуроченности кластеров локальных выборок (к районам воспроизводства или нагула), а также от-
сутствие повторяемости структуры фенонов из года в год. Наилучшим образом все эти особенности отражает концепция флюктуирующих стад (Глубоковский, Животовский, 1986). Она предполагает, что границы и численность каждого стада горбуши не являются постоянными, поскольку между ними существует флюктуирующий обмен особями. При этом в одних районах ареала вида популяционная структура может временно отвечать модели локального несмешивающегося стада (там, где интенсивность обмена особями в настоящий момент крайне мала), в других -панмиксной (где она крайне велика).
Что же касается исследования запасов горбуши, то ежегодный учет молоди осуществляется главным образом в период ската в контрольных реках. Дополнительный материал дают учетные съемки, выполняемые в отдельных прибрежных и открытых морских районах.
Несмотря на определенные успехи в изучении популяционной структуры и динамики численности азиатской горбуши, прогнозы мощности ее подходов к береговым районам промысла зачастую бывают неудачными (Шунтов и др., 1999). В связи с этим уместно вспомнить один из выводов, сделанных авторами концепции флюктуирующих стад. Они указывают, что "... прогнозирование подходов горбуши должно иметь глобальный характер, то есть следует исходить из общей оценки запасов рыб данного поколения в море и учитывать гидрологические, синоптические, антропогенные и другие факторы среды, определяющие наиболее вероятные в этот год потоки нерестовых миграций из нагульных скоплений" (Глубоковский, Животовский, 1986, с. 41). Эта задача имеет решение, если известны принципы формирования миграционных потоков.
Стремление организмов максимизировать свой энергетический выигрыш посредством оптимальной стратегии пищевого поведения имеет глубокий эволюционный смысл. Наиболее преуспевающие в этом особи лучше выполняют и репродуктивную функцию, делая больший вклад в воспроизводство. Закрепленные генетически, их навыки поддерживают жизнеспособность вида (Tounsend, Winfield, 1985; Бигон и др., 1989). Особенно важно это для короткоцикловых видов с ограниченным количеством одновременно живущих поколений, в частности для горбуши. Появление одного менее жизнеспособного поколения в популяции таких животных грозит ей резким сокращением численности.
Чтобы не истратить силы на длительное движение из нагульной в репродуктивную часть ареала, а сохранить их для подъема по рекам и успешного нереста, горбуша активно питается во время анадромной миграции. Это позволяет некоторым исследователям даже рассматривать миграцию как продолжение нагула (Шунтов, 1994а; Волков, 1996). Во всяком случае, совмещение усилий на движение и добывание пищи здесь действительно происходит. Возникая в преднерестовый период, данное обстоятельство усугубляет необходимость использования оптимальной стратегии пищевого поведения. Происходящее на этом этапе жизненного цикла созревание половых продуктов также требует определенного расхода энергии и, может быть, специфичной диеты.
Если целью оптимальной стратегии пищевого поведения является максимизация энергетического выигрыша, то задачей - оптимизация рациона, его качественное и количественное регулирование таким образом, чтобы получить наибольшую выгоду наименьшими усилиями (МасАгШиг, Pianka, 1966). При всей присущей горбуше пищевой пластичности основу ее рациона в это время составляют лишь несколько видов, набор которых изменяется от района к району. Причем соотношение этих видов в окружающей среде и в питании горбуши, как правило, различно (Волков, 1996). Проявляя избирательность, она регулирует качество рациона. Регулирование его количества достигается движением потребителя в направлении увеличения частоты встречаемости жертв. Мигрирующая горбуша обычно сосредоточивается в верхнем 30-метровом водном слое, для нее не характерна широкая амплитуда вертикальных перемещений, но горизонтальное движение далеко от прямолинейного (Oguгa, Ishida, 1995). Судя по этому, для количественного регулирования рациона она осваивает не глубины, а пространства.
Учитывая исключительное значение агрегаций пищевых организмов для формирования оптимального рациона (Ивлев, 1977; Tounsend, Winfield, 1985), можно предположить следующее. Рациональное сочетание последовательного перемещения от одной агрегации к другой с генеральным направлением движения из нагульной в репродуктивную часть ареала дает оптимальную траекторию миграции. Вероятно, стремление следовать этой траектории и лежит в основе возникновения миграционных потоков горбуши.
Ранее нами были проведены специальные исследования в тропической части открытого океана и шельфовых бореальных водах (Фещен-ко, 1994, 1996). Их результаты указывают на то, что решающее значение в образовании локальных концентраций пищевых объектов пелагических рыб имеют динамические процессы. Различного рода деформации и возмущения, пространственные размеры которых, как правило, не превышают мезомасштаба, оказывают прямое или опосредованное агрегирующее воздействие на эти организмы. Распределение таких динамических структур в поле течений, возможно, относится к факторам среды, определяющим оптимальную траекторию миграционных потоков горбуши. Попытаемся проверить это предположение.
Наличие между нагульной и репродуктивной частями ареала непрерывной цепочки динамических структур, аккумулирующих планкте-ров и мелкий нектон, могло бы существенно облегчить миграцию. Наиболее вероятно ее появление под воздействием тангенциального напряжения ветрового потока, оказывающего двоякое влияние на поверхностный водный слой. Во-первых, ветровой поток способен при определенных условиях деформировать и возмущать геострофические течения. Во-вторых, он индуцирует циркуляцию Лэнгмюра, связанную с ним строгой причинно-следственной зависимостью.
Верифицируем реалии атмосферных процессов, на фоне которых проходили анадромные миграции азиатской горбуши в период с 1993 по 1997 г. Ветры генерируются полем приземного атмосферного давления. Обычно оно и используется в промысловой океанографии для анализа, минуя расчеты ветровых полей, что значительно облегчает задачу (Федосеев, Яковлев, 1974; Берникова, 1980). В качестве исходных данных воспользуемся ежедневными картами приземного анализа ОМН) японского метеорологического агентства.
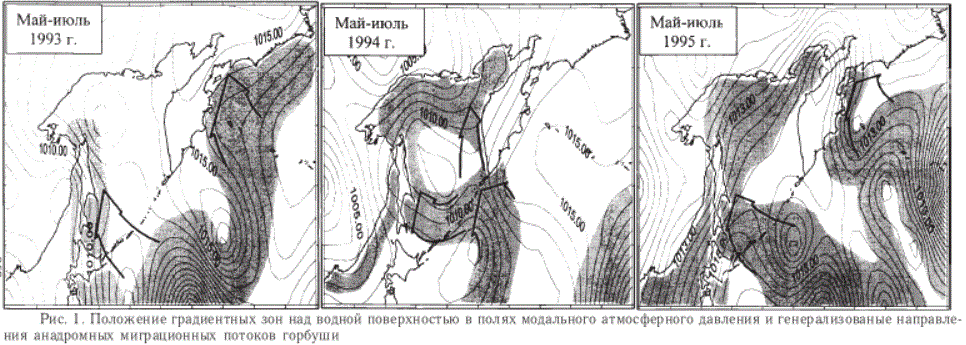
Приближение первых особей горбуши к азиатским берегам из океана отмечается обычно в конце мая - начале июня, а к августу миграционные потоки, как правило, оказываются окончательно сформированными. С учетом этих сроков ограничимся сравнением особенностей ветрового режима в мае-июле каждого из рассматриваемых лет. Давление будем снимать с синоптических карт в 33 узлах 5-градусной сетки между 40 и 65о с.ш. и 130о в.д. и 180о. В результате этого получим ежегодные ряды наблюдений в узлах сетки, которые для упрощения анализа предстоит заменить одним из стандартных показателей изменчивости ряда. При исследованиях ветрового режима такими показателями служат среднее значение и мода (Брукс, Краузерс, 1967; Пановский, Брайер, 1972). Среднее значение используется для определения "результирующего" ветра, а мода - преобладающего. Обычно к "результирующим" ветрам обращаются в теоретических исследованиях (для вычисления средней дивергенции, геострофических потоков и т.д.), для решения практических задач рекомендуется рассчитывать преобладающие ветры. Поэтому для каждого из сравниваемых периодов осуществим выборку в узлах сетки модального значения приземного атмосферного давления и проведем картирование. Выделившиеся на картах градиентные барические зоны представляют собой не что иное, как наиболее устойчивые ветровые потоки, чье воздействие на поверхностный водный слой отличалось максимальной повторяемостью.
В мае-июле 1993 г. над рассматриваемым районом наблюдалось два таких потока (рис. 1). Один из них связывал нагульную часть ареала горбуши с восточным побережьем Камчатки, другой, пересекая о. Хоккайдо и южные острова Курильской гряды, - с Сахалином.
Отслеживанием миграции лососей, идущих на нерест к азиатским берегам, в 1993 г. занималось три научно-исследовательских судна ТИН-РО. Ими контролировался район, включавший западную часть Берингова моря, юг Охотского и океанические воды вдоль Курильских островов и Камчатки. Результатом их наблюдений стала констатация наличия двух миграционных потоков горбуши, один из которых направлялся к восточному побережью Сахалина через южные курильские проливы, а другой - к восточному побережью Камчатки (Шунтов, 1994б).
Тот же период 1994 г. характеризовался наличием одного устойчивого ветрового потока, соединявшего нагульную часть ареала горбуши со средними и северными Курильскими островами (рис. 1). Здесь поток разделялся на две ветви, которые снова соединялись в непосредственной близости от места их разделения (у юго-западного побережья Камчатки). В результате ветром оказывалось воздействие практически на все периферические поверхностные воды Охотского моря.
Развитие анадромной миграции лососей 1994 г. отслеживалось двумя научно-исследовательскими судами ТИНРО, контролировавшими акваторию Охотского моря. В процессе наблюдений ими был зафиксирован массовый подход горбуши из океана к северным курильским проливам. Далее миграционный поток раздваивался: одна часть особей направлялась к юго-восточному побережью Сахалина, а другая поднималась на север вдоль Камчатки (Темных и др., 1997).
В мае-июле 1995 г. над анализируемым районом вновь господствовало несколько ветровых потоков (рис. 1). Один из них, пересекавший южные и средние Курильские острова, связывал нагульную часть ареала горбуши с восточным побережьем Сахалина, другой - с восточным побережьем Камчатки.
В этот период развитие анадромной миграции лососей отслеживалось тремя научно-исследовательскими судами ТИНРО-центра. Контролировалась акватория, включавшая западную часть Берингова моря, восток и юг Охотского и океанические воды вдоль Курильских островов и Камчатки. В ходе проведенной ими работы было зафиксировано два миграционных потока горбуши. Один пересекал южную часть Курильской гряды, направляясь к юго-восточному побережью Сахалина, другой -к восточному побережью Камчатки (Шунтов, Темных, 1996).
Отличительной чертой лета 1996 г. была аномально низкая циклоническая деятельность над северо-западной частью Тихого океана и дальневосточными морями (рис. 2). В данном случае вряд ли стоит говорить о существенном влиянии воздушных потоков на состояние динамики поверхностных вод. Можно лишь констатировать незначительное обострение барических градиентов над районом северных Курильских островов и юго-западной Камчаткой.
Наблюдения анадромной миграции лососей 1996 г., результаты которых не опубликованы, проводились двумя научно-исследовательскими судами ТИНРО-центра в южной и восточной частях Охотского моря. Вместе с учетными данными по объему вылова и пропуску на нерестилища они указывают на то, что в этом году наиболее мощный миграционный поток горбуши поднимался от северных курильских проливов вдоль западного побережья Камчатки.
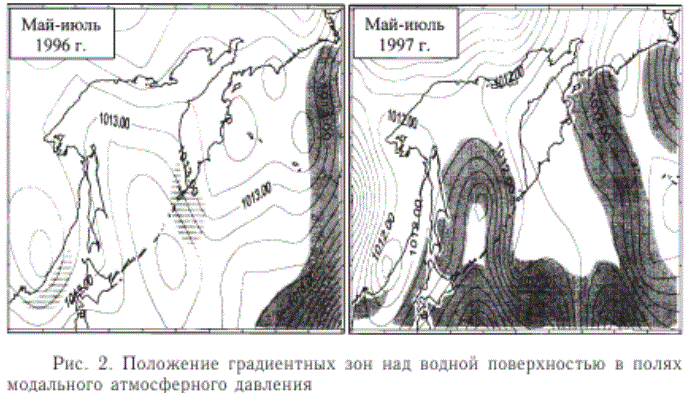
Рис. 2. Положение градиентных зон над водной поверхностью в полях модального атмосферного давления
В тот же период 1997 г. активность атмосферных процессов вернулась к обычному уровню, а над океаническими водами восточнее Японии, судя по всему, даже возросла (рис. 2). Один ветровой поток соединял нагульную часть ареала горбуши с восточным побережьем Камчатки, другой - с побережьем Сахалина. Причем траектория последнего, в отличие от прошлых лет, проходила через средние Курильские острова и центральную часть Охотского моря.
Экспедиционные морские наблюдения хода анадромной миграции лососей в 1997 г. ТИНРО-центром не проводились. Однако данные статистического учета показывают, что из 127,4 млн производителей горбуши, вошедших в Охотское море, 71 % закончили миграцию у восточного побережья Сахалина. К восточному побережью Камчатки в это время подошло рекордно высокое для анализируемых пяти лет количество -63,7 млн шт. Это означает, что в 1997 г. существовало два миграционных потока горбуши: один направлялся к восточному побережью Камчатки, другой - к восточному побережью Сахалина.
Проведенный анализ показывает значительное пространственное совпадение реально наблюдавшихся или определенных по косвенным признакам миграционных потоков горбуши и наиболее устойчивых воздушных потоков в приземном слое атмосферы. Количество отслеженных ситуаций свидетельствует о том, что это совпадение вряд ли является случайным. Поэтому предложенная трактовка механизма образования миграционного потока горбуши выглядит довольно убедительно. Но прежде чем сделать окончательные выводы, удостоверимся в том, что ее принятие не приведет к противоречию с известными фактами этологии и биологии вида.
Прежде всего обратимся к структуре миграционного потока горбуши, имеющего некоторые характерные особенности. Он отличается значительной шириной по фронту и относительной равномерностью в распределении формирующих его особей (Шунтов, 1994а). Вспомним, что между вихрями Лэнгмюра у поверхности воды образуются полосы концентрации планктона и микронектона, ориентированные параллельно друг другу и располагающиеся поперек воздушного потока (Stavn, 1971). Отсутствие фокальности при таком распределении пищи в принципе исключает проявление потребителями агрегирующего ответа. Их функциональным ответом в этом случае должно стать сравнительно равномерное распределение по всей длине полос концентрации. Там же, где встречаются агрегации пищевых объектов, горбуша не может длительно задерживаться, поскольку находится в состоянии миграции, а не нагула. Поэтому здесь она тоже не успевает создать каких-либо крупных скоплений. При этом логичное объяснение получает и другой факт - отсутствие у мигрирующей горбуши четко выраженной суточной ритмики питания (Волков, 1996): пересекая полосы концентрации пищи с гарантированной частотой и регулярностью, она имеет возможность питаться порционно, фактически не прекращая движения.
Поскольку циркуляция Лэнгмюра - явление специфичное, не имеющее аналогов среди других динамических процессов, сигнал о ее присутствии распознаваем даже в удаленных точках информационного пространства. Использование такого ориентира не должно вызывать затруднений у мигрантов.
В ежегодном чередовании количества и направления наиболее устойчивых ветровых потоков, соединяющих районы нагула и воспроизводства горбуши, явно отражается квазидвухлетняя цикличность атмосферной циркуляции. В течение примерно одного года в экваториальной стратосфере господствуют западные ветры, а в течение другого - восточные. При западных ветрах во внетропических широтах существенно возрастает интенсивность меридионального переноса, а при восточных -зонального. Это вызывает соответствующий отклик в динамике деятельного слоя океана. Средняя продолжительность одного полного цикла, включающего западную и восточную фазы, около 26 мес. Поскольку она превышает два года, время от времени происходит смена доминант (Квазидвухлетняя цикличность ..., 1971).
Как показал анализ, Курильские острова и восточный Сахалин практически всегда оказываются в зоне действия одного из воздушных потоков. Используя эффект его воздействия на поверхностный водный слой для миграции, горбуша регулярно совершала бы массовые подходы в каждый из этих районов. В результате здесь сформировались бы близкие по численности поколения четных и нечетных лет. Действительно, именно это и характерно для восточносахалинского и курильского стад в текущий период.
Совершенно иная закономерность проявляется на Камчатке. В четные годы устойчивый воздушный поток связывает районы нагула горбуши только с ее западным побережьем, а в нечетные - только с восточным. Исходя из той же логики, у западно- и восточно-камчатского стад с течением времени должны были бы закрепиться резкие различия в численности смежных поколений. Причем для первого доминирующими стали бы поколения четных лет, а для второго - нечетных. Как раз этим и характеризуется динамика численности данных стад в настоящее время.
Таким образом, рассматриваемая трактовка механизма формирования миграционных потоков горбуши дает естественное объяснение особенностям, отличающим ее популяционную организацию на первом структурном уровне. Объяснение возникновения и существования доминантных поколений, аналогично гипотезе И.Б.Бирмана (но независимо от нее), в конечном итоге основывается на воздействии квазидвухлетнего цикла солнечной активности как климатообразующего фактора.
Абсолютно устойчивых воздушных потоков в природе не существует, любой из них в той или иной мере подвержен разного рода флюк-
туациям. Благодаря наличию причинно-следственной зависимости это свойство должно передаваться и миграционному потоку горбуши. Межгодовые флюктуации мощности и направленности потоков обнаруживаются в рядах как четных, так и нечетных лет. Они изначально предопределяют изменчивость количества производителей четных или нечетных поколений, подходящих к каждому конкретному побережью, а также участков их массового подхода в пределах этого побережья. В итоге заранее обусловленным оказывается непостоянство границ и численности стад горбуши в репродуктивной части ареала.
В случаях сравнительно высокой устойчивости воздушных потоков интенсивность обмена особями между стадами окажется мала и популя-ционная структура временно будет близка модели несмешивающегося локального стада. И, наоборот, когда их устойчивость будет низкой, высокая интенсивность обмена на какое-то время приблизит популяционную структуру к панмиксной модели. Кстати, нереальность абсолютной устойчивости воздушного потока означает принципиальную невозможность даже кратковременного существования в чистом виде локальных несмешиваю-щихся стад горбуши. Его полная неустойчивость (равная повторяемость ветров разных направлений) тоже имеет малую статистическую вероятность. Поэтому непродолжительная панмиксия в отдельно взятом случае допустима скорее теоретически, чем практически.
Флюктуации воздушных потоков над нагульной частью ареала горбуши должны вызывать флюктуирующий обмен особями бассейнов Охотского и Берингова морей на начальном этапе их анадромной миграции.
Таким образом, особенности второго структурного уровня популя-ционной организации горбуши здесь тоже находят вполне убедительное объяснение.
Подводя общий итог, следует признать реалистичность рассмотренной трактовки формирования анадромного миграционного потока горбуши. Согласно ей, стремление к оптимальному питанию служит предпосылкой, наличие локальных концентраций пищевых организмов - необходимым условием, а цепное расположение таких концентраций в пространстве - достаточным условием для образования миграционного потока. В природе существует единственный процесс, следствиями которого одновременно являются и необходимое, и достаточное условие, - это возмущение поверхностного водного слоя потоком ветра. В ходе эволюции горбуша, судя по всему, приспособилась обнаруживать и использовать для своей миграции этот специфический абиотический процесс.
О.Б.Фещенко