
О СОХРАНЕНИИ ГЕНОФОНДОВ ЛОСОСЕЙ В ЛОСОСЕВЫХ РЫБОХОЗЯЙСТВЕННЫХ ЗАПОВЕДНЫХ ЗОНАХ
Л.А. Животовский
Институт общей генетики им. Н.И. Вавилова РАН (ИОГен РАН)
Введение. Прошедшие в декабре 2009 г. и апреле 2010 г. совещания по
приоритезации речных бассейнов Дальнего Востока позволили выявить узло-
вые моменты в выделении лососевых рыбохозяйственных заповедных зон
(ЛРХЗЗ). Были предложены три типа лососевых РХЗЗ (Павлов, Букварева
2009):
- ЛРХЗЗ I типа (промысловые приоритеты),
- ЛРХЗЗ II типа (рыбоводные приоритеты),
- ЛРХЗЗ III типа (приоритеты сохранения генофондов).
При выделении, организации и функционировании каждого из этих типов
ЛРХЗЗ следует сохранять популяционные генофонды лососей, что предъяв-
ляет определенные требования к природоохранным мероприятиям. Они рас-
сматриваются в настоящей заметке.
Промысловые приоритеты (РХЗЗ для сохранения промысловых по-
пуляций дикого лосося). Мероприятия в бассейне, отводимом под организа-
84
цию лососевых РХЗЗ, должны обеспечить долговременное устойчивое вос-
производство обитающих в данном водоеме популяций дикого лосося в объ-
еме рыбопромыслового значения. Одним из важных условий устойчивости
генофонда дикого лосося – это то, что вылов не должен быть селективным.
Помимо того, что в реку должно быть пропущено производителей в количе-
стве достаточном для эффективного воспроизводства добываемых видов, к
промыслу должны предъявляться обязательные требования пропускать на
нерестилища все группировки и части стада, чтобы обеспечить полное вос-
производство генофонда этого стада. Недостаточно пропустить на нерести-
лища определенное число рыб: как минимум, требование воспроизводства
генофонда достигается регулярностью пропуска лососей на нерестилища и
введением ограничений на лов вблизи устья рек ЛРХЗЗ. Почему это важно?
Из опыта разведения сельскохозяйственных животных и селекционной
практики известно, что отбор меняет наследственные биологические характе-
ристики популяции. Направленный отбор, вызванный селективным выловом,
может привести к генетическому изменению ряда важных морфо-
физиологических признаков. Хорошо известен пример многолетнего сниже-
ния размера американской горбуши вследствие использования дрифтерных
сетей с крупной ячеей, изымающих крупную рыбу и пропускающих мелкую.
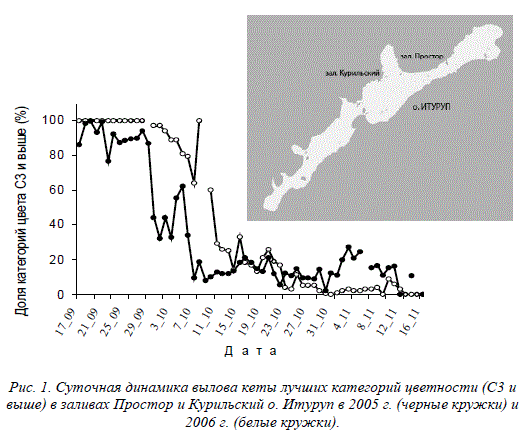
Еще один пример. На рис. 1 приведена суточная динамика высших, наиболее
ценных категорий цвета мяса в уловах кеты острова Итуруп Сахалинской
области (Животовский и др., 2010а). Ситуация такова, что до начала октября
практически вся кета здесь изымается промыслом, а это именно то время,
когда подходит рыба высших по цвету категорий (рис.1). И хотя численность
первых подходов кеты невелика, а в середине сентября – это и вовсе прилов
во время заканчивающейся горбушевой путины на Итурупе, в результате та-
кого вылова все равно идет направленный отбор против размножению рыб с
лучшим по цвету мясом, что приводит к предпочтительному воспроизводству
и увеличению в стаде кеты низших по цветности категорий.
Неравномерная по подходу промысловая нагрузка может иметь и другие
негативные генетические последствия, в том числе непропорционально
меньшее воспроизводство внутривидовых группировок лососей, что ведет к
недоиспользованию имеющихся биоресурсов.
Таким образом, промысел лосося в ЛРХЗЗ I типа обязательно следует
вести таким образом, чтобы не облавливать все начало стада и обеспечить в
течение всего нерестового хода постоянный пропуск рыбы на нерестилища.
Конечно, это требование может быть удовлетворено только при условии
проведения природоохранных мероприятий в бассейнах нерестовых рек, что-
бы зашедшие в реку производители созрели и успешно отнерестились.
Рыбоводные приоритеты (РХЗЗ для искусственного воспроизводства
малочисленных популяций лососей). Для развития сети лососевых РХЗЗ с
рыбоводными приоритетами следует предусмотреть возможность оценки
эффективности рыбоводных мероприятий. Основным показателем качества
85
работы лососевого рыбоводного завода (ЛРЗ) является коэффициент возвра-
та, прямым методом его оценки является мечение. Помимо мечения, для
оценки эффективности планируемых рыбоводных мероприятий важны дан-
ные по скату молоди и возврату производителей за ряд лет наблюдений. При
этом важно подчеркнуть, что статистический анализ данных «скат-возврат»
следует проводить индивидуально для каждого ЛРЗ, поскольку объединение
разнородных данных по разным рекам и разным рыбоводным заводам может
привести к ложным заключениям об эффективности или неэффективности
рыбоводных мероприятий (Животовский и др., 2010б). Кроме того, очевидно
важен учет ската и возврата и на реках с естественным воспроизводством, что
требует восстановления имевшихся и организации новых контрольно-
наблюдательных станций или аналогичных им структур в целях сбора све-
дений о воспроизводстве лососей в ЛРХЗЗ.
Рис. 1. Суточная динамика вылова кеты лучших категорий цветности (С3 и
выше) в заливах Простор и Курильский о. Итуруп в 2005 г. (черные кружки) и
2006 г. (белые кружки).
С позиций динамики генофонда, селективное заводское разведение лосо-
ся, как и селективный вылов, может привести к изменению наследственных
биологических характеристик воспроизводимого стада. Чтобы избежать это-
го, во-первых, следует предусмотреть, чтобы рыбоводные мероприятия были
направлены на воспроизводство всех группировок и частей стада (Алтухов и
др., 1997; Животовский, 2006). Во-вторых, следует иметь в виду возможные
взаимодействия заводских и диких популяций. Действительно, имеются кос-
венные данные, что заводской лосось имеет худшую приспособленность к
условиям дикого местообитания и после первого поколения воспроизводства
на нерестилищах их потомки все еще имеют более низкую приспособлен-
ность, чем дикие рыбы (Araki et al., 2009). Поэтому теоретически не исклю-
чено ухудшение генофонда диких популяций лососей при смешении с заво-
дскими рыбами, однако такое ухудшение может произойти только при замет-
ном различии генофондов диких и заводских популяций. Для видов с воз-
можно высоким уровнем стреинга, таких как горбуша, генофонды территори-
ально близких диких и заводских популяций могут оказаться практически
идентичными. В таком случае генетического ухудшения диких рыб при их
смешении с заводскими не ожидается. При высоком уровне стреинга, кроме
чисто генетических взаимодействий, могут быть экологические взаимодейст-
вия заводских и диких рыб: в виде конкуренции за пищу – особенно в ранний
морской период, если выпуск заводской молоди и скат с нерестилищ прихо-
дятся на одни и те же сроки; в виде разной интенсивности пресса хищников и
непропорционального вылова заводских и диких группировок; в виде вытес-
нения диких производителей заводскими на нерестилищах и пр. Поэтому в
любом случае важным является вопрос оценки уровня стреинга у тихоокеан-
ских лососей, в том числе заводских рыб в дикие популяции и диких рыб в
заводские популяции.
В статье Спрингмейера и др. (2007) дан подход к количественной оценке
состояния речных бассейнов на примере о. Сахалин по наличию в них диких
популяций лососевых рыб. В этой работе авторы оценивают величину стре-
инга по расстоянию от ЛРЗ, предполагая, что уровни стреинга одинаковы для
горбуши и кеты. Однако не исключено, что стреинг и генные потоки у азиат-
ской горбуши значительно превышают таковые у других видов лососей
Дальнего Востока, в частности – у кеты (Глубоковский, Животовский, 1986),
поэтому ее генетическая дифференциация на порядок ниже, чем у кеты (см.
Животовский и др., 1989; Noll et al., 2001; Рубцова и др., 2009). Следователь-
но, необходимым является детальное маркирование молоди горбуши и кеты,
а также оценка их популяционно-генетической структуры с целью выявления
подразделенности нерестовых группировок и взаимодействия диких и заво-
дских популяций этих видов на российском Дальнем Востоке, стратегии вос-
производства горбуши (Животовский и др., 2010в).
Проблема селективности лова и разведения. Селективность лова и се-
лективность разведения приводят к схожим генетическим последствиям: уст-
ранению определенной части производителей от размножения, то есть к от-
бору. При промысле и разведении возможно изменение генофонда популя-
ции через направленный отбор. Поэтому уделим вопросам направленного
отбора еще внимания.
Из общей селекционной теории (Фолконер, 1985) следует, что отбор спо-
87
собен значительно изменить биологические характеристики стада по селек-
тируемым признакам и взаимосвязанным с ними особенностями, что под-
тверждается обширными экспериментальными данными, в том числе на ры-
бах (например, Kincaid et al., 1977; Campton and Gall, 1988). Направленный
отбор (в форме селективного вылова и/или селективного воспроизводства) в
течение нескольких поколений может заметно изменить процент быстро и
медленно созревающих рыб, сдвинуть время нерестового подхода, изменить
другие важные признаки. Например, в начале рунного хода основная часть
подходящей рыбы изымается промыслом, что фактически оборачивается ин-
тенсивным отбором против ранней рыбы (Животовский и др., 2010а), по-
скольку сроки возврата наследственно обусловлены (Smoker et al., 1998;
McGregor et al., 1999; Sato et al., 2000). Далее, закладка икры на инкубацию
производится лишь при массовом подходе зрелых производителей: чем
меньше времени рыба выдерживается в ловушках до созревания, тем удобнее
работать рыбоводам. Тем самым рыбоводный процесс также преобразует
стадо, ведя еще одну форму искусственного отбора – на наследственное за-
крепление в стаде быстро созревающих производителей. В частности, за счет
такого отбора увеличивается доля рыб, рано развивающих признаки брачной
окраски, примером чего является заводская кета юго-западного Сахалина
(Рухлов, 1980).
Преодолеть негативные последствия селективного вылова и селективного
разведения лосося можно лишь, обеспечивая постоянный пропуск в реки и
воспроизводство как дикой, так и заводской рыбы в течение всего нерестово-
го хода. Что касается заводской рыбы, то в воспроизводстве необходимо
обеспечить равное участие ранних производителей, подходящих к забойкам с
менее развитыми гонадами, для чего следует реконструировать забойки и
построить бассейны с регулируемой подачей воды для длительного выдержи-
вания незрелых производителей.
Селективный промысел и селективное воспроизводство, помимо отбора
против ранней рыбы в пределах одной популяции, могут приводить к элими-
нации разных популяционных компонентов в сложноорганизованной попу-
ляции, в том числе внутривидовых форм с разным временем хода, разными
темпами созревания, с разной экологией. Поэтому промысел и разведение
должно вестись так, чтобы минимизировать сопутствующие негативные се-
лективные эффекты.
Приоритеты сохранение генофонда (РХЗЗ для сохранения биоразно-
образия лососей). Генетический статус сохраняемых форм. Говоря о под-
держании биоразнообразия, следует иметь в виду важность сохранения раз-
нообразия не только на уровне видов, но и на уровне внутривидовых группи-
ровок. К ним относятся сезонные и другие экологические формы, включая
различные жизненные стратегии. Важность их заключается в том, что все
вместе они обеспечивают приспособление популяций к варьирующим усло-
виям среды, в том числе в местах морского нагула (Geiger et al., 1997). Необ-
88
ходимое требование при этом – чтобы группировки были генетически иден-
тифицируемы. Устойчиво воспроизводимые широко расселенные формы,
такие как сезонные расы, различаются генетически (Алтухов и др., 1997). В
то же время следует иметь в виду возможность существования эндемичных
группировок, которые также могут быть выделены как элементы биоразнооб-
разия. Однако выделения их по морфо-физиологическим критериям как объ-
екта сохранения недостаточно, поскольку различия между ними могут быть
вызваны средовыми факторами и не иметь генетической основы – их приро-
доохранный статус обязательно должен быть подтвержден генетическими
критериями.
В качестве примера приведем озерную форму кеты, встреченную только
на Южных Курилах и выделенную Иванковым (1985), которая отличается по
форме и размерам тела от речного экотипа и нерестится на выходах грунто-
вых вод озерных нерестилищ (Каев и др. 1996). Ее генетическое отличие от
речной кеты было нами установлено через анализ микросателлитных марке-
ров (Каев и др. 2008), почему и была предложена нами в качестве объекта
внимания в работе по сертификации морского промысла лососей на Итурупе
(Отчеты ООО Гидростроя, 2006-2008). Рис. 2 показывает, что озерная кета,
нерестящаяся в различных водоемах о. Итурупа, образует единый генетиче-
ский кластер. Поэтому включение озерной кеты в перечень уникальных
группировок лососевых рыб при выделении ЛРХЗЗ является экологически и
генетически обоснованным. Более того, уникальные группировки других ви-
дов лососей, такие как сахалинский таймень или нерка о. Итуруп, могут
иметь генетически различные формы из-за длительной репродуктивной изо-
ляции от других популяций и потому также требуют тщательного генетико-
экологического исследования для выявления их генетической уникальности в
природоохранных целях – как виды, встречающиеся в приловах добываемых
тихоокеанских лососей.
Оценка биоразнообразия. Важным аспектом сравнительной оценки во-
доемов в целях сохранения генофондов является выбор количественной меры
биоразнообразия. Рассмотрим такой условный пример. Если прежде в водоем
заходило 1 тыс. экз. симы, 9 тыс. кеты и 40 тыс. горбуши, а затем числен-
ность горбуши возросла за счет кеты и в стало 1 тыс. экз. симы, 1 тыс. кеты и
48 тыс. горбуши, то как считать – биоразнообразие осталось тем же или оно
уменьшилось? С одной стороны, число видов не изменилось, а с другой сто-
роны данное сообщество стало более моновидовым – в основном представ-
ленным горбушей. Обе точки зрения имеют свое обоснование, и обе следует
учитывать.
Обычной мерой биоразнообразия является видовое богатство (richness), то
есть число различных видов (число генетически различных форм) в данном
местообитании или биоценозе. Эта мера дает важную информацию о широте
видового состава вне зависимости от того редкие это формы или частые. С
другой стороны, с позиций теоретической экологии, редкие виды и формы
89
имеют меньшую ценность для функционирования экосистемы и тогда боль-
шее значение в оценке биоразнообразия приобретают доминирующие виды,
то есть биоразнообразие оценивается с учетом относительного представи-
тельства разных видов. Для учета этого обстоятельства предложены различ-
ные показатели разнообразия (diversity – Krebs, 1999). Один из них – индекс
Симпсона
 2 , где i p – это частота вида (генетической формы) под
2 , где i p – это частота вида (генетической формы) подномером i среди наличествующих k видов. Этот индекс оценивает, в основ-
ном, только доминирующие виды, поскольку возведение в квадрат частоты
i p резко уменьшает вклад в величину индекса, вносимый редкими формами.
Например, виды с частотами встречаемости 0.5 и 0.1 вносят вклады в величи-
ну индекса равные 0.25 и 0.01, соответственно, то есть вклад частой формы в
величину индекса в 25 раз превышает вклад более редкой формы, хотя их
частоты различаются только в 5 раз. Таким образом, индекс Симпсона пре-
увеличивает вклад часто встречающихся форм и преуменьшает вклад более
редких форм в общую оценку биоразнообразия.
Для объективной количественной оценки биоразнообразия могут быть
два подхода. Один из них – это совместное применение двух индексов (видо-
вого богатства и индекса Симпсона): вместе они позволяют учесть разные
стороны биоразнообразия, поскольку первый учитывает даже единично
встречающиеся формы (виды), а второй – преимущественно только домини-
рующие, часто встречающиеся формы. Другой подход – это использование
такой единой меры разнообразия, которая пропорционально учитывает как
частые, так и редкие формы и виды. Одна из таких мер – это показатель μ,
первоначально предложенный для оценки аллельного разнообразия популя-
ций:
 (Животовский 1980). Показатель μ подчеркивает при-
(Животовский 1980). Показатель μ подчеркивает при-сутствие редких форм, поскольку, в сравнении с индексом Симпсона, он
нормализует вклад редких форм в количественную величину показателя раз-
нообразия. Например, для рассмотренного выше условного примера с часто-
тами встречаемости частой и редкой форм, равными 0.5 и 0.1, вклад частой
формы в величину показателя μ превышает вклад редкой формы только в два
раза, в отличие от непропорционального превышения (в данном примере – в
25 раз) при использовании индекса Симпсона.
Популяционные базы ДНК-данных. Организация системы ЛРХЗЗ не-
обходимо приведет к системе мониторинга популяций лососей, одним из
компонентов которого является создание баз данных по генетике стад лосо-
сей в зоне ЛРХЗЗ, в первую очередь – по ДНК-маркерам. Такие базы данных
необходимы как для выделения популяционных группировок и оценки их
генетической структуры, так и проведения прикладных исследований по сер-
90
тификации промысла, определения популяционных компонентов в смешан-
ных уловах, выявления популяционной принадлежности рыб в спорных си-
туациях, решения других проблем ДНК-идентификации лососевых рыб. Нам
сейчас создается глобальная база ДНК-данных по кете (Животовский и др.
2008, Афанасьев и др. 2010), которая сейчас включает более 200 выборок со
всех регионов российского Дальнего Востока и уже использована для реше-
ния генетических проблем экологической сертификации морского промысла
на о. Итуруп (Животовский и др. 2010г); она может быть использована в ка-
честве реперной при генетическом мониторинге стад кеты РХЗЗ. Подобные
базы данных начинают разрабатываться и для других видов лососевых рыб.
Выводы.
1. Для ЛРХЗЗ первого типа (с приоритетом промысла дикого лосося), не-
обходимо исключить селективность лова, для чего обеспечить пропуск про-
изводителей на нерестилища в течение всего нерестового хода.
2. Для ЛРХЗЗ второго типа (с рыбоводными приоритетами) необходимо
исключить селективность воспроизводства, закладывая на инкубацию икру от
производителей в течение всего нерестового хода, а также воспроизводя ге-
нетически различные группировки (скажем, речную и озерную формы кеты)
без смешения их друг с другом.
3. При оценке биоразнообразия применительно к выбору ЛРХЗЗ третьего
типа (с приоритетом сохранения генофондов) использовать индексы биораз-
нообразия, позволяющие учитывать в равной мере как часто встречающиеся,
так и редкие формы и виды.
4. Статус внутривидовой формы следует придавать только при условии
ее наследственного своеобразия – на основе значимых, устойчивых по годам
и водоемам, генетических отличий этой формы от других популяционных
группировок данного вида.
5. Необходимо определить интенсивность и дальность стреинга у тихо-
океанских лососей в целях оценки взаимодействия между различными груп-
пировками, в т.ч. между заводскими и природными популяциями лососей.– в
первую очередь, у горбуши и кеты – как путем мечения, так и через сравне-
ние их генетических параметров.
6. Необходимо создать популяционные базы ДНК-данных по лососевым
рыбам в ЛРХЗЗ в целях выделения популяционных группировок, проведения
исследований по сертификации промысла, определения популяционных ком-
понентов в смешанных уловах, ДНК-идентификации в спорных ситуациях.
Благодарности.
Настоящая работа была поддержана грантом программы Президиума
РАН «Биологическое разнообразие – Генофонды».