
Кета (ONCORHYNCHUS KETA (WALBAUM)) в Дальневосточных морях - биологическая характеристика вида 2. Питание и трофические связи кеты в эпипелагиали дальневосточных морей и сопредельных водах Тихого океана
Питанию тихоокеанских лососей в морской период жизни посвящено значительное количество работ (Андриевская, 1968; Heard, 1991; Salo, 1991; Волков, 1996; King, Beamish, 2000; и др.). Однако в течение многих лет при изучении питания лососей в море в распоряжении исследователей были в основном пробы, полученные при дрифтерном лове этих рыб, которые не позволяли получить количественную информацию о питании и редко охватывали суточную циклику (Волков, 1996). Возможности изучения питания лососей несоизмеримо возросли в 1980-е гг., когда был освоен их лов при помощи больших пелагических тралов, позволяющих облавливать лососей любых размеров и в любое время суток.
Неоднократно отмечалось (Шунтов и др., 1992; Лапко, 1996), что анализ влияния отдельных факторов обычно не дает возможности заметить четкие закономерности в распределении и миграциях тихоокеанских лососей. В естественных условиях действие разных факторов бывает комплексным. Комплексной проблемой, в частности, являются питание и трофические отношения, определяющие не только многие стороны экологии рыб, но и структуру ихтиоценов. Исследование трофических связей массовых видов лососей важно с позиций оценки их кормовой обеспеченности и роли в трофической структуре экосистемы. По отношению ко многим представителям планктона и нектона кета проявляет себя как хищник, в то же время являясь кормовым объектом для хищных рыб, морских млекопитающих и птиц. С другой стороны, поведение и распределение рыб, в том числе лососей, зависят от наличия и количества объектов питания, а также от их доступности, что в свою очередь связано с океанологическими условиями.
В последние годы на динамику популяций тихоокеанских лососей все большее влияние оказывает их искусственное разведение (Beamish et al., 1997; Heard, 1998). Наиболее значительным результатом современного лососевого рыбоводства является создание Японией крупнейшего по биомассе индустриального ста-
да кеты. К 1990-м гг. биомасса возврата японской кеты превысила 200 тыс. т и колеблется от 143 до 228 тыс. т (Kaeriyama, 1998). Такого количества рыбы на ее традиционных пастбищах никогда не было. Это не могло не изменить экологической ситуации в океане. В этих условиях стали актуальными вопросы обеспеченности кормовыми ресурсами тихоокеанских лососей, их пищевых и пространственных отношений в период сезонного нагула в дальневосточных морях и открытых водах Северной Пацифики.
Помимо этого, произошедшие климато-океанологические перестройки в пелагических сообществах дальневосточных морей, одним из отражений которых явилось " исчезновение" иваси, снижение численности минтая, доминировавшего во многих эпипелагических ихтиоценах (Шунтов и др., 1997; Шунтов, 1998), вызывают несомненный интерес к вопросу, изменились ли в новых условиях трофические связи тихоокеанских лососей, в том числе кеты?
Начиная с 1986 г. и по настоящее время при изучении питания лососей в экспедициях ТИНРО применяются методические подходы, которые по своей сути являются новаторскими (Чучукало и др., 1994; Волков, 1996). Метод экспресс-обработки качественного и количественного состава пищи рыб, применяемый в комплексных экспедициях специалистами лаборатории планктона дальневосточных морей, позволяет обрабатывать и анализировать данные непосредственно в море. Это дает возможность проведения их оперативного анализа, а не только в ретроспективном аспекте.
Пробы для изучения питания кеты отбирали из каждого результативного траления в количестве до 25 экз. по размерным группам — 11-20, 21-30, 31-40, 41-50, 51-60, 61-70 см. Все желудки обрабатывались сразу, без предварительной фиксации. Определяли тотальную массу интегральной пробы, степень пере-вариваемости (по 5-балльной шкале), видовой состав пищи, значимость массовых видов, общие и частные индексы наполнения желудков. В статье приведены данные, полученные при исследовании питания 5623 экз. кеты.
Суточные рационы рассчитывали по методу А.В.Коган (1963) и Ю.Г.Юро-вицкого (1962). Более подробно методики сбора и обработки данных изложены в "Руководстве по изучению питания рыб" (1986), а также в работах сотрудников ТИНРО-центра (Волков, 1996; Чучукало, 1996; и др.).
Первичный анализ проб по питанию кеты и расчет суточных рационов был выполнен сотрудниками лаборатории планктона дальневосточных морей ТИНРО-центра д.б.н. А.Ф.Волковым, к.б.н. В.И.Чучукало, к.б.н. К.М.Горба-тенко, А.Я.Ефимкиным, Н.А.Кузнецовой и др. Автор считает своим долгом выразить благодарность сотрудникам этой лаборатории за любезно предоставленные материалы.
Расчеты потребления кетой кормовых объектов определялась по следующей формуле:
B = Bsp ■ CP ■ n ■ 0,01,
где В — потребляемая биомасса, тыс. т; Bsp — биомасса кеты определенной размерной группы, тыс. т; СР — суточный рацион, %; n — количество суток.
Доля каждой группы гидробионтов в общем потреблении определялась исходя из их массовой доли в содержимом желудков кеты.
Использованы схемы биостатистического районирования, предложенные В.П.Шунтовым (1986, 1988) на начальных этапах экосистемных исследований. В частности, по этим районам рассчитывались средние величины рационов и потребление корма кетой.
Питание кеты в период посткатадромных миграций
В ранний морской период жизни молодь кеты является в основном планк-тофагом и поедает значительное количество различных животных, населяющих поверхностные воды морских прибрежий (Карпенко, 1998; Иванков и др., 1999).
Период прибрежного обитания молоди кеты можно разделить на два этапа, каждый из которых характеризуется своеобразными условиями нагула. Первый этап протекает на прибрежных мелководьях и в бухтах, второй — на открытой акватории заливов. Особенностью первого этапа нагула является переход значительной части молоди кеты на активное внешнее питание (Каев, 1983).
После миграций в эстуарии рек и с выходом в литоральные и открытые воды бухт и заливов спектр питания молоди всех видов тихоокеанских лососей значительно расширяется за счет планктонных ракообразных и личинок рыб (Карпенко, 1998). Переход на питание морскими организмами происходит сразу, как только молодь скатывается в предустьевые пространства (Иванков и др., 1999). Если в реках интенсивность питания молоди значительно колеблется, то в море, где количество пищи больше, интенсивность питания возрастает и изменяется меньше, чем в реках. Интенсивность питания в предустьевых пространствах сразу после ската молоди кеты из рек достигает 250-300 %о (Фроленко, 1970; Кун, 1986).
Состав пищи молоди кеты в значительной степени определяется особенностями района обитания и его кормовой базой: численностью и доступностью кормовых организмов, а также наличием пищевых конкурентов. Часто молодь двух наиболее массовых видов — горбуши и кеты — образует скопления в одних и тех же местах (Карпенко, 1998). При этом пищевое сходство рассматриваемых видов весьма велико и увеличивается по мере удаления от берега. И после рассредоточения по акватории заливов наблюдается значительное сходство в составе пищи молоди кеты и горбуши (СП-коэффициент достигает 76,992,9 %), что связано не столько со сходством в экологии этих видов на данном этапе, сколько с обилием кормовых организмов, составляющих основу питания кеты и горбуши (Андриевская, 1968). Это также свидетельствует о существовании на межвидовом уровне адаптации к условиям обитания на начальных этапах жизни в море (Каев, Чупахин, 1986).
В зависимости от района обитания пищевой спектр молоди кеты в прибрежье могут слагать до 50 и более видов разных таксономических групп животных (Андриевская, 1968; Каев и др., 1993). При этом зачастую для нее более важное значение имеют размеры животных, а не видовая принадлежность. Поэтому сеголетки кеты чаще избирают мелких ракообразных и личинок насекомых, составляющих основу их пищи в разных регионах. В связи с относительно длительным периодом обитания в непосредственной близости от берега в пище молоди кеты продолжительное время встречаются животные пресноводной и солоноватоводной фаун (Гриценко и др., 1987; Карпенко, 1998). Для прибрежья юго-восточной части Татарского пролива известен случай питания молоди кеты практически исключительно воздушными насекомыми (в основном Díptera) , которые относились от берега сильным ветром на расстояние одной и более миль (Фроленко, 1965). Выносимые из рек воздушные насекомые и их личинки являются основной (от 38,0 до 89,3 % по массе) пищей молоди кеты и в литоральной зоне Карагинского залива (Карпенко, 1998). В.Н.Иванков с соавторами (1999) также указывает на значительную долю (50-80 % по массе) насекомых в желудках молоди кеты, отловленной вблизи устий рек западного побережья Сахалина.
Помимо этой группы пищевых организмов, в южных районах у азиатского побережья в пище молоди кеты доминируют гарпактициды, амфиподы, гамма-риды и мелкие копеподы (Каев, Чупахин, 1986; Иванков и др., 1999), а в северных — кумовые раки и гаммариды (Фроленко, 1970; Карпенко, 1998; Путивкин, 1999). Подросшая и откочевавшая мористее молодь кеты переходит на питание каланидами, амфиподами, гипериидами, эвфаузиидами и другими морскими формами зоопланктона (Бирман, 1969; Каев и др., 1993).
В целом по типу питания кета и горбуша преимущественно характеризуются как планктонофаги, чавыча, кижуч и сима — как хищники-нектонофаги, а нерка — как вид со смешанным типом питания (Горбатенко, Ч учукало, 1989; Волков, 1994; Волков и др., 1997; Глебов, 2000). С другой стороны, тихоокеанские лососи отличаются высокой пластичностью питания и способностью потреблять кормовые объекты с различной скоростью переваривания (Горбатенко, 1996; Дулепо-ва, 1998).
Сразу после выхода из прибрежья посткатадромная молодь кеты начинает потреблять как зоопланктон, так и молодь различных видов нектона (рыб и кальмаров). Условия среды обитания в каждом году обусловливают специфику спектра потребления кетой кормовых организмов. Хотя в последующий морской период у кеты зачастую прослеживается избирательность в питании, тем не менее в зависимости от сезона, района и года исследований спектр питания кеты одной возрастной группы может сильно изменяться (Горбатенко, Чучукало, 1989; Тутубалин, Чучукало, 1992; Волков и др., 1997; и др.).
По данным наших исследований, молодь кеты (средние размеры тела 14,516,9 см), вышедшая в открытые морские районы южной части Охотского моря в июле 1993 г., предпочитала питаться гипериидами. Их доля в ее рационе составляла 74,5 % при среднем индексе наполнения желудков (ИНЖ) — 160,8 %оо. Среди прочих пищевых компонентов в это время в рационе кеты были заметны эвфаузииды (13,1 %) и птероподы (9,3 %). О преобладании в пище молоди кеты гипериид в центральных районах Охотского моря сообщалось и ранее (Андриевская, 1968). По А.Ф.Волкову (1996), питание сеголеток кеты в Охотском море (прикурильские воды и район Центральной котловины) преимущественно гипериидами (Themisto japónica, Primno abissalis) и эвфаузиидами (Thysanoessa longipes , Euphausia pacifica) в целом является типичным. Несмотря на изменения их значимости в рационах кеты в различных районах прослеживается четкая избирательность по отношению к гипериидам: повсеместно гиперииды потребляются сеголетками кеты в большем количестве, чем их содержится в планктоне (Волков, 1996). Здесь, вероятно, проявляется отбор, связанный с размерами кормовых организмов, их агрегированностью и ценностью в пищевом отношении: как известно (Кизеветтер, 1954), в летнее время гиперииды отличаются от других форм кормового зоопланктона наиболее высокой калорийностью.
В начале августа 1994 г. в прикурильских водах Охотского моря молодь кеты также отдавала предпочтение гипериидам — 80,0 %. Вторым по значимости пищевым объектом для нее здесь были копеподы — 17,5 %. Общий индекс наполнения желудков кеты в это время составлял 189 %о. Как в 1993 г., так и в 1994 г. спектр питания кеты, размеры которой не превышали 20 см (средняя длина 17,9-19,8 см), был не широк: его слагали 3-4 группы пищевых организмов.
Летом 1997 г. (10 июля — 6 августа) кета, пойманная в зал. Шелихова, также питалась гипериидами (Themisto libellula) и молодью тихоокеанской сельди. Причем в одном желудке пищевой комок полностью слагали гиперииды (ИНЖ 256 %оо), а другие сеголетки (11 экз.) питались исключительно молодью сельди (ИНЖ 353 %оо). Также нектоном питалась в 1997 г. молодь кеты (35 экз.) в североохотском и западнокамчатском районах. В первом из них в пище кеты исключительно преобладала молодь минтая, во втором — мальки песчанки (60 % по массе) и молодь минтая (40 %).
Как известно (Шунтов и др., 1993), шельф западнокамчатского и североохо-томорского побережий Охотского моря является местом крупного воспроизводства минтая, мальки которого, наряду с молодью сельди, песчанки и других рыб, составляют важную часть рациона тихоокеанских лососей. В то же время преобладание рыбной составляющей в рационе кеты не характерно для ее питания в глубоководных и присваловых районах (Горбатенко, Чучукало, 1989; Волков, 1996).
Выше мы уже упоминали о равномерном и повсеместном распространении молоди кеты вдоль западнокамчатского шельфа в летний период. Очевидно, сеголетки кеты не стремятся покидать воды шельфа, а активно здесь нагуливаются.
Такая стратегия питания (концентрация на небольшой акватории, богатой высококалорийными кормами), видимо, оправданна и успешно реализуется не только кетой, но и другими лососями, например кижучем и чавычей (Глебов, 2000).
Наиболее представительные данные по питанию сеголеток кеты в северной и центральной частях Охотского моря были получены осенью 1994 г. В общей сложности было обработано 403 желудка кеты размерных групп 11-20 и 21-30 см. Питание сеголеток кеты в это время было схожим с летним периодом наблюдений, однако наряду с этим заметны и некоторые особенности. Копе-поды (главным образом Neocalanus plumchrus) встречались в пище кеты постоянно, но не играли главной роли: лишь над впадиной ТИНРО их доля составила 27,7 %. Гиперииды (Themisto japónica, Th. pacifica и Th. libellula) в питании молоди имели более существенное значение (30,0-60,4 %) повсюду, кроме за-паднокамчатского биостатистического района 7 и ямско-тауйского района 2. В первом из этих районов главными в питании кеты были птероподы (53,7 %) и эвфаузииды (23,8 %), а во втором — в желудке единственного экземпляра кеты были обнаружены практически исключительно (95 %) медузы. Заметной была доля птеропод и в зал. Шелихова: 26,2 % — у кеты, размеры которой не превышали 20 см, и 55,4 % — у кеты размерной группы 21-30 см. Нектон (в основном молодь минтая) имел существенное значение в питании кеты в районе банки Кашеварова (37,0 %) и в зал. Шелихова (17,7 %). К этому добавим, что накормленность кеты, как и летом 1997 г., была наибольшей при преобладании в составе пищи нектона (до 379 %о) или птеропод (267-348 %о). Это, на наш взгляд, связано с тем, что при равных объемах в содержимом желудков масса зоопланктона меньше, чем нектона, и соответственно ниже ИНЖ вследствие меньшей плотности пищевого комка. Кроме того, скорость переваривания зоопланктона выше. Возможно, это одна из причин, благодаря которой доля зоопланктона в массе содержимого желудка редко бывает такой высокой, как нектона.
Предпочтение молоди нектона кетой в отдельных районах, как и в предыдущих случаях, объяснимо теми же причинами: молодь минтая, как, впрочем, и других видов рыб и кальмаров, в период ее концентрации в районах воспроизводства (шельф западной Камчатки в летний и зал. Шелихова в осенний период) и в районах нагула (банка Кашеварова) является массовым, доступным и энергетически выгодным объектом питания. В значительной степени это связано со стайным поведением молоди рыб и кальмаров и их более крупными размерами, по сравнению с зоопланктоном. Это облегчает их обнаружение и уменьшает энергозатраты хищником на поиск. При рассеивании скоплений подросшей молоди нектона требуются более значительные энергетические траты на питание таким кормом, и поэтому становится более выгодно питаться зоопланктоном. В районах, где концентрации молоди нектона в эпипелагиали незначительны (южная часть Охотского моря и тихоокеанские воды Курильских островов), подобная стратегия питания кеты нами замечена не была.
Крылоногие моллюски — еще одна группа жертв, которая избиралась кетой в районах их высоких концентраций. Как известно (Чучукало, Напазаков, 1998), нерест этих планктеров происходит в шельфовой зоне. Там же во все сезоны наблюдаются и максимальные биомассы крылоногих. В Охотском море такими районами являются зал. Шелихова и прилегающие к нему акватории. Здесь биомасса крылоногих моллюсков (главным образом Limacina helicina) составляет более 20 мг/м3, а в открытых водах она, как правило, в пять—десять раз ниже (Чучукало, Напазаков, 1998). Именно в этих районах в питании кеты птеропо-ды имеют первостепенное значение. В районах же их низких концентраций кета переключается на питание иными объектами.
В южной части Охотского моря в сентябре 1991 г. исследования проводились в открытых водах глубоководной части моря (биостатистический район 9). В это время крылоногие моллюски не входили в состав доминирующей группы
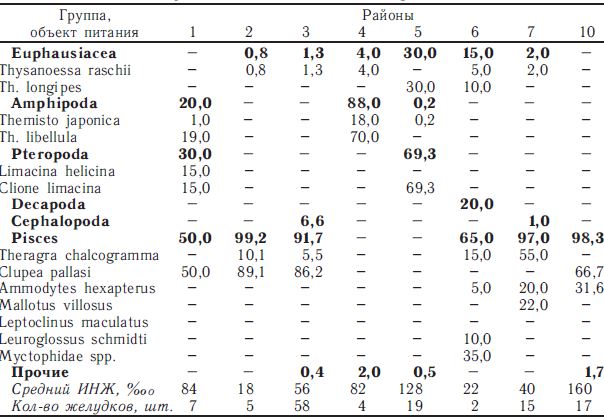
в питании сеголеток кеты. Однако в октябре—ноябре в прикурильских водах они уже занимали одно из первых мест в ее рационе, хотя в планктонном сообществе эта группа макропланктона не была лидером (биомасса крылоногих в период исследований составляла 0,5-2,3 мг/м3). Схожая ситуация наблюдалась и в тихоокеанских районах 8 и 10 (табл. 1). Объяснить это можно, с одной стороны, тем, что сеголетки кеты избирали эту группу планктона, а с другой — особенностью распределения крылоногих моллюсков, которые держатся тонким слоем у поверхности и плохо облавливаются планктонной сетью (Волков, 1996).
Таблица 1
Состав пищи сеголеток кеты (22-28 см) в южной части Охотского моря и сопредельных водах Тихого океана в сентябре—октябре 1991 г., %
 в южной части Охотского моря и сопредельных водах Тихого океана в сентябре—октябре 1991 г., %")
* Суммарно с Neocalanus flemingeri.
Помимо птеропод важной составляющей рациона сеголеток кеты, вне зависимости от соотношения их биомасс в планктонном сообществе, являются гипе-рииды и эвфаузииды. Так, значимость массовых групп крупной фракции зоопланктона в питании сеголеток кеты и их соотношение в планктоне осенью 1991 г. было следующим: в трех районах Охотского моря эвфаузииды составляли 10,0-19,5-34,9 % суммарного рациона сеголеток кеты, а в тихоокеанских водах — соответственно 6,3-11,4-13,9 %. В то же время биомасса этой группы зоопланктона в южной части моря была 106-158-7 мг/м3, а с тихоокеанской стороны Курильских островов — соответственно 35-514-69 мг/м3.
Схожими были соотношения потребления кетой гипериид и их учтенных биомасс: в океане 10,7-37,3-84,5 % и 4-21-17 мг/м3 и соответственно 9,028,3-62,0 % и 38-74-22 мг/м3 — в Охотском море. Из приведенных соотношений заметно, что интенсивность питания кеты этими группами макропланктона не соответствует (в отличие от ее питания молодью нектона) формуле — "много корма — интенсивнее потребление". По-видимому, представители зоопланктона (эвфаузииды, гиперииды, копеподы и пр.) для нагуливающейся кеты являются лишь составляющими "общего набора пищевых компонентов", среди которых она не всегда отдает предпочтение одному—двум ее представителям.
В осенний и зимний периоды 1994 и 1995 гг. в южной части Охотского моря сеголетки кеты, достигшие средних размеров 23,3-27,4 см, питались гипе-риидами (главным образом это были Themisto pacifica и Primno abissalis), эвфау-
Таблица 2
Состав пищи сеголеток кеты в южной части Охотского моря и сопредельных водах Тихого океана в ноябре—декабре 1994 г., %

Таблица 3
Состав пищи сеголеток кеты в южной части Охотского моря и сопредельных водах Тихого океана 13.11-02.12.1995 г., %

* Суммарно с Neocalanus flemingeri.
зиидами (Euphausia pacifica) и реже хетог-натами и оболочниками (табл. 2-3). Молодь рыб в желудках кеты отмечалась только в тихоокеанских водах: 3,0-8,2 % в районе 7 и 12,2 % — в районе 8. Интенсивность питания сеголеток кеты в южной части Охотского моря и в сопредельных водах Тихого океана была низкой и редко превышала 70 % о (см. табл. 1-2).
Таким образом, осенью—зимой в южной части Охотского моря и прилегающих тихоокеанских водах рацион кеты ниже по калорийности (по сравнению с северными шельфовыми районами), и здесь ниже интенсивность ее питания. С другой стороны, как мы отмечали выше, именно в южной части Охотского моря создается комплекс наиболее благоприятных гидрологических условий для нагула сеголеток лососей (главным образом горбуши и кеты) вплоть до глубокой зимы.
О суточной ритмике питания сеголеток кеты в южной части Охотского моря в прикурильских водах Тихого океана, необходимой для расчетов суточных пищевых рационов, можно судить из данных А.Ф.Волкова (1996). Сеголетки кеты начинают активно питаться и одновременно переваривать пищу после 9-10 ч утра. Максимальное наполнение желудков наблюдается с 11-13 до 15-17 ч дня и далее плавно уменьшается, достигая минимума к 6-8 ч утра (Волков, 1996).
Из этого следует, что основное потребление пищи происходит в светлое время, а максимальный пик пищевой активности в течение суток только один.
В дальнейшем, с перераспределением кеты в районы открытых океанических вод, изменяется и спектр ее питания. Исследования, проведенные в феврале—марте 1991 г. в открытых водах Тихого океана, показали, что по мере продвижения кеты на восток в ее рационе доля гипериид снижается с 47,7 до 7,4 %, копепод — с 10,9 до 5,9 %. В то же время происходит увеличение значимости хетогнат (Sagitta elegans) c 30,1 до 67,8 %. При этом суммарные значения ИНЖ составляли соответственно 91,8 %оо в западной и 89,4 %оо в восточной частях района исследований. Несмотря на то что интенсивность питания кеты в обоих районах была сопоставима, в последнем энергетическая ценность пищи, несомненно, была ниже.
Материалы по питанию сеголеток кеты в Беринговом море и прилегающих водах восточной Камчатки менее представительны, чем для Охотского моря. При летних исследованиях 1991-1995 гг. (июнь—июль) молодь кеты в этих районах если и встречалась, то единично. Осенью в открытых водах Берингова моря (сентябрь—октябрь 1986 г.) спектр питания сеголеток кеты слагали только три группы зоопланктона: гиперииды (52,8 %), эвфаузииды (46,2 %) и пте-роподы (1,0 %). В зал. Карагинском (район 10) доля эвфаузиид в питании кеты была больше — 87,5 %, а гиперииды при этом составляли 12,5 % массы пищевого комка. Схожим было питание посткатадромной кеты в олюторско-наваринском районе Берингова моря и в октябре—ноябре 1987 и 1988 гг. Основу питания составляли эвфаузииды (главным образом Thysanoessa inermis) и гиперииды (Th. pacifica). В шельфовых районах юго-западной части Берингова моря сеголетки преимущественно потребляли эвфаузиид (до 91 % рациона). Доля же гипериид достигала лишь 6,0 %.
В сентябре—октябре 2000 г. питание кеты размерных групп 11-20 и 2130 см (средняя длина 17,2 и 21,2 см) в юго-западной части Берингова моря было более разнообразным (табл. 4). Однако и в этом случае среди прочих доминировали 2-3 компонента питания. У кеты до 20 см это были Eucalanus bungii (66,6 % массы пищевого комка) и Sagitta elegans (19,2 %), а кета, размеры которой превышали 20 см, более интенсивно потребляла гипериид — Themisto pacifica (32,2 %), ойкоплевр — Oikopleura labradoriensis (29,2 %) и копепод — Eucalanus bungii (21,5 %).
Следует обратить внимание на полное отсутствие нектона в пище сеголеток кеты в западной части Берингова моря, что, возможно, связано с небольшим количеством проб (желудков), собранных здесь в летний и осенний периоды. С другой стороны, частота встречаемости молоди нектона в содержимом желудков кеты в значительной мере соответствует их процентной доле среди потенциальных жертв в море (как, например, в различных районах Охотского моря). В сентябре до 61-75 % всех сеголеток минтая в западной части Берингова моря концентрируется в прол. Литке (Соболевский и др., 1991) и в Карагинском заливе. Кроме "местной" молоди минтая здесь происходит постоянное пополнение с нерестилищ, находящихся северо-восточнее, на шельфе Олюторского залива (Радченко и др., 1990). Однако в этих районах кета предпочитает питаться зоопланктоном.
В водах восточной Камчатки состав питания сеголеток кеты схож с таковым в Беринговом море и также не отличается большим разнообразием. В ноябре—декабре 1986 и 1991 гг. в глубоководных тихоокеанских районах 5 и 6 посткатадромная молодь кеты предпочитала гипериид (соответственно 54,3 и 46,7 %). В 1986 г. большую долю в ее рационе составляли сагитты (Sagitta elegans) — 28,0 %, а осенью 1991 г. — гребневики (Beroe cucumis) — 38,6 %.
Таблица 4
Состав пищевого рациона сеголеток кеты в западной части Берингова моря 26.09-19.10.2000 г., %

Доля эвфаузиид здесь в оба года не превышала 5-7 % и была сопоставима с ойкоплеврами — 5,16,8 %. Следует отметить небольшую долю в пище сеголеток кеты кальмаров (5,7 %) и молоди рыб (0,1 %). Накормленность кеты в осенний период в тихоокеанских водах была относительно низкой — 63,9-95,6 %оо.
Подытоживая вышеизложенное, следует отметить, что таксономический состав рациона молоди кеты в водах дальневосточных морей и сопредельных водах океана в целом остается постоянным, хотя процентное соотношение отдельных групп кормовых организмов в нем варьирует. При питании сеголетки кеты отдают предпочтение гипериидам и реже эв-фаузиидам. С другой стороны, спектр питания сеголеток кеты изменяется по биотопам пелагиали (шельфовые или глубоководные районы). При этом прослеживаются существенные различия в качественном составе и интенсивности питания, которые находятся в зависимости от количества доступных жертв и их калорийности. Так, в шельфовых и мелководных районах Охотского моря (западная Камчатка, зал. Шелихова, банка Кашеварова) рацион посткатадромной кеты существенно пополняется за счет молоди рыб и птеропод, но в центральных глубоководных районах моря, где концентрации молоди нектона и птеропод низкие, она питается исключительно зоопланктоном. Рацион сеголеток кеты после их миграции в тихоокеанские воды (зима—весна) складывается за счет зоопланктона.
Питание кеты старших возрастных групп в период нагула и преданадромных миграций
Основные материалы по питанию неполовозрелой и созревающей кеты в дальневосточных морях и сопредельных водах океана были собраны в летних экспедициях 1991-1997 гг. и осенних 1986-1987 и 1999-2000 гг. Результаты были осреднены по биостатистическим районам. Накормленность и состав пищи кеты в разных районах и даже на разных станциях одного района сильно различаются, но тем не менее осреднение по районам позволило выявить как избирательность, так и региональные особенности питания и состава кормовой базы кеты.
Трофические связи кеты старших возрастных групп в планктоне и нектоне Берингова моря и сопредельных водах восточной Камчатки довольно разнообразны и по сравнению с другими видами лососей имеют свою специфику (Старовой-тов, 1999). Качественный состав пищи нагуливающейся и преданадромной кеты в Беринговом море в летний период складывался из 8-9 групп планктона и нектона. Ч асто в питании (как, впрочем, и в планктоне) доминируют одна—две, реже — три группы организмов. В пище кеты старших возрастов наиболее типичны птероподы и эвфаузииды. Из прочих можно отметить аппендикулярий, гребневиков и реже гипериид. При этом, так же как у кеты младших возрастов (сеголеток), видовой состав питания состоит из сравнительно небольшого числа видов.
Выше мы упоминали о дискретности распределения разных возрастных когорт нагульной кеты, что во многом определяет состав их пищевого рациона (Горбатенко, Чучукало, 1989). Тем не менее заметны и некоторые региональные различия. Например, в северной части Берингова моря в отдельные годы в пищевом спектре кеты доминируют аппендикулярии (Oikopleuгa spp.). Так, осенью 1987 г. они составляли до 95 % весовой доли рациона кеты в устьевой части Анадырского залива (Радченко, 1994). Однако идентификация аппендику-лярий в пищевом комке кеты вызывает определенные трудности. В частности, по мнению А.Ф.Волкова (1996), сходный с ними вид имеют полупереваренные птероподы СНопе Нтаста, что может привести к завышению значимости в питании аппендикулярий и занижению значимости птеропод.
По данным наших исследований летом 1991 г., в пяти мелководных районах — 11 (Берингово море), 1-4 (районы восточной Камчатки) — и в центральном глубоководном районе 12 до половины (41,4-58,5 %) рациона кеты размерной группы 51-60 см приходилось на аппендикулярий. В тихоокеанских водах Камчатки по мере удаления в океан и в более северном беринговоморс-ком районе 8 значимость этой группы в питании заметно снижалась (рис. 1). В глубоководной части последнего (северо-восточнее мыса Олюторского) кета питалась в основном эвфаузиидами (55,4 % ее рациона). При этом при относительно малой доле эвфаузиевых в планктоне (по биомассе — 31 мг/м3) они занимали основное место в рационе кеты (рис. 1). В других районах доля эвфа-узиид в желудках кеты была не больше пятой части.
Из других групп пищевых объектов в водах восточной Камчатки выделялись птероподы (до 22,4-23,1 %) и гребневики (до 16,8-19,9 %). Заметными здесь были и кальмары, особенно в мористой части (до 28,8 %), в то время как в Беринговом море эта группа нектона полностью отсутствовала в желудках кеты. Интенсивность питания кеты была наибольшей в Беринговом море (рис. 1).
Летом 1992 г. в мелководном районе 11 в пище кеты доминировали птероподы (СНопе Нтаста), аппендикулярии и рыба (преимущественно молодь минтая и мойвы), при относительно невысокой накормленности (55,5 %оо). В зал. Кара-гинском приоритеты в питании кеты сместились в пользу нектона (минтай и личинки камбал) — 37,6 % и аппендикулярий — 25,5 %. Для двух других районов (9 и 12) было характерно большее разнообразие спектров питания кеты, чем в предыдущих. Здесь присутствовала кета трех размерных групп (3140, 41-50 и 51-60 см), при этом основная масса в уловах приходилась на неполовозрелых рыб.
В Командорской котловине (район 12) кета отдавала предпочтение птеропо-дам, эвфаузиидам и частично гипериидам (особенно более крупные рыбы). Встречаемость этих групп зоопланктона в желудках кеты распределялась следующим образом: птероподы — 32,5-26,1-38,3 %, эвфаузииды — 15,8-19,8-15,6 %, соответственно для размерных групп 31-40, 41-50 и 51-60 см. Значимость в питании кеты молоди рыб заметно снижалась с размерами тела (14,8-4,8-3,2 %), напротив, амфиподы (гиперииды) чаще отмечались в рационе более крупных рыб (0,5-17,1-11,6 %). Помимо прочих, в пище кеты, размеры которой составляли 31-40 см, доля Sagitta е^а^ достигала 13 %.
В целом же в различных районах кета в основном потребляла наиболее массовые и, вероятно, наиболее доступные пищевые ресурсы. В северо-западной части моря летом 1991 и 1992 гг. таким ресурсом являлись аппендикулярии, составлявшие до 55 % состава мезозоопланктона (0,4-0,6 мг/м3), а в Карагинском заливе летом 1992 г. — сеголетки минтая (73,5 % от 70,5 тыс. т ихтиомассы).
Летом 1993 и 1995 гг. спектры питания кеты выглядели иначе. Прежде всего следует отметить, что, несмотря на межгодовую и региональную динамику в предпочтении того или иного объекта питания, наблюдалась общая "птеропод-ная" избирательность в питании кеты практически всех размеров. Суммарная доля двух видов крылоногих моллюсков (СНопе Нтаста и Ытаста helicina) в рационе кеты зачастую превышала 50 % (рис. 2, табл. 5-6).
 кеты размерной группы 51 см и более (кольцевые диаграммы) и состав крупной фракции")
Рис. 1. Спектр питания (% массы пищи) кеты размерной группы 51 см и более (кольцевые диаграммы) и состав крупной фракции зоопланктона (%) в верхней эпипе-лагиали (круговые диаграммы) в западной части Берингова моря и сопредельных водах Тихого океана 22.06.-10.07.1991 г.: 1 — рыба, 2 — эвфаузииды, 3 — ойкоплевры, 4 — птероподы, 5 — гребневики, 6 — кальмары, 7 — копеподы, 8 — сагитты, 9 — гиперииды, 10 — прочие. Цифры в центре циклограммы — средние значения индекса наполнения желудка (%оо), цифры в квадратах — номера районов
Fig. 1. Diet composition of chum salmon (percent from food weight) with fork length 51 cm and more (ring chart) and composition of zooplankton in epi pelagic layer (circle chart) in western part of Bering Sea and the adjacent waters of Pacific Ocean in 22.06.10.07.1991: 1 — fish; 2 — euphausiids; 3 — tunicates; 4 — pteropods; 5 — gelatinous zooplankton; 6 — squids; 7 — copepods; 8 — chaetognaths; 9 — amphipods; 10 — others. Number in center of ring chart — average index of stomach fullness (%оо), number in square — number of region
Эвфаузиевые отмечены в питании кеты различных размеров. Но в целом их доля редко превышала 20 % суммарного рациона всех групп планктона и нектона. Для кеты размерной группы 41-50 см наиболее весомое значение в питании кеты эвфаузииды имели летом 1993 г. в 5-м океаническом районе, достигая одной трети от величины суммарного рациона (рис. 2). Гиперииды отмечены в питании кеты той же размерной группы и более мелкой в океанических 5 и 6-м районах (соответственно 17,2 и 27,0 %). Аппендикулярии были обнаружены только в желудках крупных рыб в прибрежных 1-4-м районах, где их доля достигала 14,5 % (рис. 2).
Вероятно, качественные различия рационов кеты в рассматриваемые годы (1991-1993 и 1995) повлияли на их величину. Рацион половозрелой и неполовозрелой кеты в водах западной части Берингова моря летом 1992, 1993 и 1995 гг. не превышал 2,0-2,5 % массы тела, а в 1991 г. составил 4,6 %. При этом в 1991 г. в нем было гораздо больше аппендикулярий, что снизило калорийность рациона кеты. Например, калорийность относительного рациона в 1995 г. была выше, чем в 1991, в 1,6 раза (Дулепова, 1998).
 в верхней эпипелагиали западной части Берингова моря и в")
Рис. 2. Спектр питания кеты и состав крупной фракции зоопланктона (%) в верхней эпипелагиали западной части Берингова моря и в тихоокеанских водах Камчатки 16.06.-09.07.1993 г.: 1 — рыба, 2 — эвфаузииды, 3 — ойкоплевры, 4 — птероподы, 5 — кальмары, 6 — амфиподы, 7 — сагитты, 8 — копеподы, 9 — прочие. Одинарные циклограммы — размерная группа 51-60 см, двойные — 41-50 см (внутреннее кольцо) и 51-60 (внешнее кольцо), тройные — 31-40 см (внутреннее кольцо), 41-50 (среднее кольцо) и 51-60 см (внешнее кольцо). Цифрами обозначены средние для всех размерных групп кеты ИНЖ (%оо)
Fig. 2. Diet composition of chum salmon and zooplankton composition in upper layer (circle chart) in western part of Bering Sea and the Kamchatka's adjacent waters of Pacific Ocean in 16.06.-09.07.1993: 1 — fish; 2 — euphausiids; 3 — tunicates; 4 — pteropods; 5 — squids; 6 — amphipods; 7 — chaetognaths; 8 — copepods; 9 — others. Single circle charts represent chum salmon with fork length 51-60 cm, double circle charts represent 41-50 cm (inner circle) and 51-60 cm (outer circle) chum salmon , triple circle charts represent 3140 cm (inner circle), 41-50 cm (medium circle) and 51-60 cm (outer circle) chum salmon. Numbers are the average stomach fullness indices of all size groups of chum salmon (%о)
Из нектона в питании нагуливающейся и созревающей кеты летом 1993 и 1995 гг. выделялись головоногие моллюски. Так, молодь кальмаров интенсивно потреблялась ею в западной части Алеутской котловины (район 8). Здесь же и в прилегающем глубоководном районе 12 в рационе кеты была высокой и доля молоди рыб (рис. 2, табл. 5).
Данные осенних исследований 1986-1987 гг. показывают, что к этому времени в рационе кеты возрастает доля эвфаузиид, гипериид, личинок десятиногих раков. На карагинском шельфе созревающие особи в этот же период интенсивно питались сеголетками минтая (72 % рациона), реже — молодью мойвы и птероподами. Различия в составе пищи нагуливающейся и созревающей кеты, вероятно, определялись особенностями вертикального распределения, так как у этих групп рыб даже в одном районе горизонты обитания могут существенно различаться (Ogura, Ishida, 1995; Walker et al., 2000).
В Беринговом море в сентябре—октябре 2000 г. кета также интенсивно потребляла молодь сельди, песчанки, минтая и миктофид (табл. 7). При этом
Таблица 5
Состав пищевого рациона кеты в различных районах Берингова моря в июне—июле 1995 г., %

* Суммарно с Neocalanus flemingeri.
Таблица 6
Состав пищевого рациона кеты в тихоокеанских водах восточной Камчатки в июле 1995 г., %
* Суммарно с Neocalanus flemingeri.
максимальной была встречаемость первых двух видов жертв у кеты, чьи размеры превышали 51 см. Вторым по значимости объектом в питании кеты практически всех размерных групп оказались аппендикулярии (Oikopleura labradoriensis).
Таблица 7
Состав пищевого рациона кеты разных размерных групп в Беринговом море 26.09-19.10.2000 г., %

По осредненным данным В.И.Радченко (1994), основанным на материалах исследования питания кеты в западной части Берингова моря в 19871992 гг., крылоногие моллюски составляют не менее 36,0 % рациона данного лосося. Не менее 0,7 % слагают аппендикулярии, 14,1 % приходится на долю трудно идентифицируемых остатков первых двух групп, 19,0 % составляют эвфаузииды, 10,2 % — гиперииды, 7,9 % — рыбы, 5,5 % — личинки креветок и крабов, 3,0 % — копеподы, 2,4 % — молодь кальмаров, 1,0 % — сагитты.
Наши оценки среднего количественного рациона нагуливающейся и созревающей кеты (по данным 1993 и 1995 гг.) для Берингова моря и вод восточной Камчатки показывают несколько отличные результаты. Так, доля крылоногих моллюсков в ее рационе была 54,8 и 50,5 %, соответственно в море и тихоокеан-
ских водах. На эвфаузиид приходилось 17,5 и 10,4 %, на гипериид — 7,3 и 6,2 %. Молодь рыб составила 4,5 и 5,0 %, кальмаров — 5,4 и 2,6 %. Копеподы занимали 0,7 % рациона кеты в Беринговом море и 10,2 % — в водах восточной Камчатки, а аппендикулярии соответственно 6,5 и 9,6 %.
Несомненно, осреднение количественного состава рациона весьма приблизительно отражает реальную картину и в каждый конкретный год он может существенно изменяться в зависимости от ситуаций в планктонном сообществе. Тем не менее наиболее важным следствием нам представляется вывод о "птероподной" специализации в питании кеты в Беринговом море и сопредельных водах океана в конце 1980 — начале 1990-х гг. Вследствие этого большую долю аппендикулярий в составе пищевого комка в отдельных районах, вероятно, следует соотносить не с одной, а с двумя группами зоопланктона (аппендикулярии и птероподы) и с их осредненными долями в другие годы.
Что касается прочих групп жертв, необходимо отметить низкую долю в рационе кеты гипериид и копепод, первые являются основой питания сеголеток кеты, а вторые в изобилии встречаются в планктоне и в питании других пелагических рыб, в частности минтая (Шунтов и др., 1993).
В Охотском море и сопредельных водах Тихого океана в период летних исследований спектры питания нагуливающейся и созревающей кеты более разнообразны, чем в Беринговом море и тихоокеанских водах восточной Камчатки. Тем не менее доминирующую группу пищевых объектов зачастую составляют те же виды, что и в Беринговом море.
В прикурильских водах Тихого океана (районы 7, 8 и 9) в июле 1993 г. кета размерных групп 51-60 и 61-70 см питалась преимущественно птероподами (рис. 3). Второй по значимости группой были эвфаузииды — 25,0-31,8 %. Кета крупнее 31 см питалась эвфаузиидами и в Охотском море. Их доля в ее рационе во всех районах исследований превышала 50 % (рис. 3). Отметим, что эта группа была заметна и в планктонном сообществе обследованного региона, только в 10-м районе и в прикурильском районе 13а эвфаузииды не доминировали в макропланктоне. Аналогичная ситуация наблюдалась и летом 1995 г. Несмотря на очень низкую биомассу этой группы в планктонном сообществе, кета активно потребляла эвфаузиид в прикурильских водах.
Прочие группы жертв в пище кеты были менее заметны. Гиперииды отмечались в ее желудках чаще в Охотском море, в частности в районе 9 — 16,0-29,4-6,2 % — и в районе 10 — 9,2-15,0-15,6 %, — соответственно у кеты размерных групп 31-40, 41-50, 51-60 см. В прикамчатском районе 8 в питании рыб крупнее 51 см гиперииды составили 6,8 %. Как и эвфаузииды, эта группа макропланктона в планктонном сообществе не выделялась (особенно в районах 9 и 10), однако была заметна в рационах кеты. Напротив, копеподы, несмотря на их заметную роль в планктоне, составляли лишь 0,1-0,3 % среди прочих объектов ее питания.
Нектоном питается в большей степени нагуливающаяся кета, чем созревающие рыбы. Молодь рыб отмечалась в рационе кеты в прибрежном восточно-сахалинском районе 10 (11,1 и 6,4 % соответственно для неполовозрелых и половозрелых рыб) и в районе 9 (12,2 и 3,6 %). В этих же районах и прибрежных водах западной Камчатки кета потребляла также молодь кальмаров. Доля головоногих моллюсков была большей у крупноразмерных рыб (51 см и более) в прибрежных районах: район 8 — 12,0 % и район 10 — 12,3 % (рис. 3).
Нектонная составляющая в рационе кеты преобладала и в июне—июле 1995 и 1997 гг. Хотя летом 1995 г. зоопланктон в ее питании присутствовал, однако основную роль играли представители нектона. В глубоководной котловине южной части моря питание кеты базировалось практически полностью (82100 %) на рыбах (миктофиды, серебрянка, молодь терпуга и анчоуса). В прикурильском районе (13б) рыбная составляющая содержимого желудков нагульной
 кеты в различных районах южной части Охотского моря и в тихоокеанских водах Курильских островов 13-28.07.1993 г. Условные обозначения как на рис. 2")
Рис. 3. Спектр питания (%) кеты в различных районах южной части Охотского моря и в тихоокеанских водах Курильских островов 13-28.07.1993 г. Условные обозначения как на рис. 2
Fig. 3. Diet composition of chum salmon in southern part of Okhotsk Sea and the Pacific waters of Kuril Islands in 13-28.07.1993. Designations follow those in Fig. 2
кеты также оказалась высокой — 43,5 %. И летом 1997 г. в южной части Охотского моря молодь одноперого терпуга и миктофиды интенсивно потреблялись кетой. Доля этих рыб в ее питании достигала 77,1-83,1 %.
В противоположность неполовозрелым рыбам, в рационе преданадром-ной кеты часто доминируют неценные в пищевом отношении виды. Так, в прикурильских районах 13а и 13б и в тихоокеанских 9 и 10 летом 1993 г. в питании кеты оказалась весомой роль Beroe cucumis. Значимость гребневиков в ее рационе достигала 80 %. При этом средний индекс наполнения желудков был самым высоким в тихоокеанских водах (ИНЖ 101 %оо), при подавляющем преобладании гребневиков. На наш взгляд, эта особенность питания (желетелыми пищевыми компонентами — медузами, гребневиками и реже сагиттами (В.И.Чучукало, личное сообщение)) присуща созревающей кете в период преданадромных миграций. Подобный тип питания был характерен для половозрелых рыб в прикурильских и тихоокеанских районах в августе—сентябре 1994 г. С охотоморской стороны южных Курильских островов доля медуз в рационе половозрелых рыб достигала 56-60 % массы пищевого комка, а с тихоокеанской (район 10) — 31,4-39,6 %. В последнем из указанных районов, помимо этого, в рационе половозрелых рыб в значительных количествах (26,7-64 %) отмечались сальпы. В прилегающих к рассматриваемым районах 12 (Охотское море) и 9 (Тихий океан) роль желете-лых в пище кеты была заметно ниже.
В северо-восточной части Охотского моря в июне—июле 1995 г. приоритетными группами в пище кеты также были эвфаузииды и птероподы (рис. 4).
Несмотря на небольшую значимость в планктоне (кроме западнокамчатского района 8, где эвфаузииды занимали по биомассе первое место — 1234 мг/м3, или 59 % всех групп макропланктона), рацион кеты, в особенности крупноразмерных половозрелых рыб, складывался практически за счет эвфаузиид (районы 69) и птеропод. Последние активно потреблялись в северных мелководных районах (зал. Шелихова и прилегающие воды), где их биомасса, в отличие от эвфау-зиид, летом значительно выше, чем на западнокамчатском шельфе и в открытых водах: до 20 мг/м3 и более — Ытаста helicina и до 10 мг/м3 и более — СНопе Нтаста (Чучукало, Напазаков, 1998).
Рис. 4. Спектр питания кеты (%) в водах западной Камчатки летом 1995 г.: 1 — рыба, 2 — эвфаузииды, 3 — ойкоплевры, 4 — амфиподы, 5 — птероподы, 6 — кальмары, 7 — кишечнополостные, 8 — сагитты, 9 — копеподы, 10 — прочие. Внутреннее кольцо циклограммы — размерная группа 41-50 см, внешнее — 51-60 см, в центре — средние значения индекса наполнения желудка (%оо)
Fig. 4. Diet composition of chum salmon in northwest part of Okhotsk Sea in summer of 1995: 1 — fish; 2 — euphausiids; 3 — tunicates; 4 — amphipods; 5 — pteropods; 6 — squids; 7 — gelatinous zooplankton; 8 — chaetognaths; 9 — copepods; 10 — others. The inner circle of circle chart represents 41-50 cm chum salmon and outer circle represents 51-60 cm chum salmon. Numbers are the average stomach fullness indices of all size groups of chum salmon (% о)
Среди прочих в питании кеты здесь отмечались молодь рыб (главным образом минтая) и головоногих моллюсков. Интересно отметить, что потребление кетой старших возрастов нектона происходило в тех же районах, где этой группой питались и сеголетки кеты (выше мы упоминали об этом). Однако доля относительно подвижных организмов (молодь рыб и кальмаров) в рационе старших возрастных когорт оказалась ниже, чем у сеголеток кеты, — 5,4-20,9 % (рис. 4).
В северной части Охотского моря летом 1997 г. основу питания кеты размерной группы 41-50 см во 2 и 3-м районах также составлял нектон, а в 5 и 6-м районах — зоопланктон (в основном эвфаузииды и гиперииды). Крупная кета (51-60 и 61-70 см) практически повсеместно потребляла нектон (табл. 8-9).
Таблица 8
Состав питания кеты размерной группы 51-60 см в северной части Охотского моря в июне—августе 1997 г., %

Таким образом, как и у сеголеток, в северной части моря в летний период в питании кеты старших возрастов зачастую преобладает нектон, в то время как в южной рацион кеты в большей степени складывается из представителей зоопланктона.
К осени в северо-восточной части Охотского моря нектон практически исчезает из состава рационов кеты. В октябре—ноябре 1994 г. основу питания неполовозрелых рыб (размеры 31-40 и 41-50 см) составляли только две группы зоопланктона — гиперииды (73,5-80,0 %) в районе 6 и эвфаузииды (100 %) в районе 7. В это время половозрелые рыбы (размеры 51-60 и 6170 см), пойманные в зал. Шелихова, потребляли только птеропод (81,0 и 70,4 %) и медуз (19,0 и 29,6 %). Напротив, в ионо-кашеваровском районе (северо-западная часть моря) рацион кеты складывался практически исключительно из молоди минтая (95 % массы пищевого комка), при этом ИНЖ достигал 139 %оо.
В южной части Охотского моря в осенний период рыбная составляющая рационов кеты более значительна в прикурильских водах как со стороны Охотского моря, так и с тихоокеанской стороны. В октябре 1999 г. кета питалась
Состав пищи кеты размерной группы 61-70 см в северной части Охотского моря в июне—августе 1997 г., %
Diet composition of chum salmon with fork length 61-70 cm in northern part of Okhotsk Sea in June—August of 1997, %
Таблица 9
здесь рыбами-мигрантами из мезопелагиали (главным образом Stenobrachшs leucopsarus и S. паппосЫг). В южной глубоководной котловине моря рацион нагульной кеты на треть складывался из анчоуса (Engraulis ]арошс^). О преимущественном питании мезопелагическими рыбами в тихоокеанских водах Курильских островов и восточной Камчатки сообщалось и ранее. Так, по данным Н.В.Кловач с соавторами (1996) (питание кеты исследовалось из уловов дрифтерных сетей), встречаемость рыб в желудках кеты летом 1994 и 1995 гг. составляла 54,1 %, ракообразных — 33,7 %, молоди кальмаров — 12,2 %.
Схожие материалы из уловов дрифтерных сетей были получены и нами. Столь существенная роль нектона (а именно мезопелагических рыб) в питании кеты, на наш взгляд, связана с тем, что все пробы из дрифтерных сетей были взяты от рыб, пойманных в ночные часы, когда последние активно питаются поднимающимися к поверхности миктофидами, серебрянкой и другими рыбами мезопелагиали. В то же время материалы по питанию, собранные при траловом лове лососей (пробы от рыб, пойманных в разное время суток), показывают более широкие спектры питания кеты.
К концу осени (ноябрь 1995 г.) в южной части Охотского моря единичные экземпляры половозрелой кеты питались эвфаузиидами и гребневиками. В тихоокеанских водах (районы 9 и 10) в состав пищи этой группы рыб (размеры 5160 см) входила молодь японского анчоуса (18,4-64,6 %), а в районе 7 питание половозрелой кеты складывалось практически исключительно из медуз (93,7 %).
В дополнение приведем информацию об интенсивности питания кеты в течение суток. О суточной ритмике питания кеты старших возрастных групп в различных районах можно судить по осредненным данным летних исследований. Практически во всех районах интенсивность питания кеты была максимальной лишь единожды за сутки. В водах западной Камчатки наблюдалось два незначительных всплеска — между 8 и 12, и 16 и 20 ч. В водах южной части Охотского моря и восточной Камчатки кета наиболее интенсивно питалась с 12 до 16 ч. В Беринговом море интенсивность ее питания была максимальна в утренние часы — с 8 до 12, а в тихоокеанских водах Курильских островов — в вечерние, с 16 до 20.
Суммируя приведенные данные по питанию кеты старших возрастных когорт в Охотском море и сопредельных водах Тихого океана, можно отметить, что стратегия ее питания не всегда соответствует типичным планктофагам. Так же как и у сеголеток, в рационах нагуливающейся и созревающей кеты в районах концентраций нектона (зал. Шелихова и прилегающие воды, ионо-кашеваровс-кий район, прикурильские воды) в летнее время преобладает нектон, среди которого основную роль играют молодь пелагических рыб и мезопелагические мигранты. Типичная для Берингова моря "птероподная специализация" в Охотском море у кеты проявляется в местах воспроизводства и повышенных концентраций крылоногих моллюсков. Из представителей зоопланктона ею явно избираются эвфаузииды. Еще одна особенность питания кеты в Охотском море — большая доля в содержимом желудков созревающей (половозрелой) кеты же-летелых организмов, — вероятно, связана с физиологическим состоянием этой группы рыб на данном этапе жизни.
Трофические связи кеты в нектонных сообществах Берингова моря и водах восточной Камчатки
Как следует из описанного выше, наиболее полные данные по питанию кеты в юго-западной части Берингова моря и водах восточной Камчатки были получены в первой половине 1990-х гг. и в 2000 г. По материалам 1993, 1995 и 2000 гг. в Беринговом море суточное потребление зоопланктона половозрелой и неполовозрелой кетой составило 0,9, 1,2 и 0,3 тыс. т. В водах восточной Камчатки в летний период 1992 и 1995 гг. кета потребляла 0,2 и 1,3 тыс. т корма в сутки.
Исходя из 90-суточного рациона (время, в течение которого основная масса кеты нагуливается в этих регионах), величина потребления кетой зоопланктона и нектона в западной части Берингова моря в 1993 и 1995 гг. составляла соответственно 80,14 и 107,84 тыс. т, а в тихоокеанских водах Камчатки — 21,28 и 113,49 тыс. т. За два осенних месяца 2000 г. потребление кетой корма составило 17,2 тыс. т. При этом на долю ее сеголеток пришлось 0,15 тыс. т, а на неполовозрелых рыб — 16,7 тыс. т.
В летний период, как видно из данных табл. 10, оценки запаса двух групп макропланктона — эвфаузиид и амфипод (являющихся одними из основных компонентов в питании тихоокеанских лососей) — в Беринговом море и в тихоокеанских водах Камчатки были ниже в 1995 г. по сравнению с 1993 г. Биомасса копепод, напротив, возросла в 1995 г. соответственно в 1,7 и в 2,9 раз (Волков, 1996). Сезонное потребление кетой эвфаузиид и амфипод было на два—три порядка ниже их запаса в верхней эпипелагиали, а потребление копепод было и вовсе незначительным (табл. 10). В противоположность, сезонное потребление птеропод превосходило запас этой группы планктона в Беринговом море, а в тихоокеанских водах Камчатки биомасса птеропод не превысила 2,5 величины сезонных рационов кеты (табл. 10).
Как отмечалось выше, в рассматриваемых регионах крылоногие моллюски повсеместно являются излюбленной пищей кеты и активно ею потребляются, что дает основание говорить о напряженном состоянии запаса этого кормового объекта. Правда, нужно иметь в виду, что из-за особенностей распределения крылоногих моллюсков, которые держатся узкими слоями, зачастую не удается получить реальных оценок их обилия.
Таблица 10
Запас (верхняя эпипелагиаль) и сезонное потребление основных групп зоопланктона в Беринговом море и тихоокеанских водах Камчатки в летний период 1993 и 1995 гг., тыс. т
 и сезонное потребление основных групп зоопланктона в Беринговом море и тихоокеанских водах Камчатки в летний период 1993 и 1995 гг., тыс. т")
Осреднение данных, полученных в юго-западной части Берингова моря в первой половине 1990-х гг., позволяет представить обобщенную схему трофических связей кеты старших возрастов в эпипелагиали моря в период ее летнего нагула следующим образом. Максимальные величины потребления наблюдались в отношении птеропод и эвфаузиид (рис. 5). Значительно меньшую, но все же заметную роль играют представители нектона (рыбы — 3,7 и головоногие — 3,0 тыс. т) и гиперииды (2,7 тыс. т). Наименьшая интенсивность потребления наблюдается по отношению к хетогнатам (0,2 тыс. т). Одна особь кеты в период летнего нагула в юго-западной части Берингова моря в среднем потребляет 4,3 кг корма, что соответствует 48 г за одни сутки.
 кетой кормовых организмов при среднем для первой половины 1990-х гг. уровне биомассы кеты")
Рис. 5. Трофические связи кеты в эпипелагиали юго-западной части Берингова моря в период летнего нагула. Цифрами обозначено выедание (тыс. т) кетой кормовых организмов при среднем для первой половины 1990-х гг. уровне биомассы кеты
Fig. 5. Trophic linkages of chum salmon in epipelagic layer of southwest Bering Sea during the summer period. Numbers designate consumption of prey organisms (in
thousands of metric tons) by chum salmon. An average biomass of chum salmon and its daily rations during the first half of 1990s were used
Эвфаузиид ы 19.5
Изложенное выше может быть дополнено данными по выеданию основных групп планктона в западной части Берингова моря наиболее массовыми представителями эпипелагического нектона. Как известно (Дулепова, 1998), состав пищевых спектров различных видов тихоокеанских лососей в Беринговом море в качественном отношении на уровне таксономических групп практически совпадает. Основные различия сводятся к специализации в питании и предпочтении той или иной группы пищевых организмов. Так, для горбуши свойственна эвфаузиидная "специализация", хотя этот вид наиболее пластичен в питании по сравнению с другими видами тихоокеанских лососей (Дулепова, 1998). Кета предпочитает крылоногих моллюсков, реже эвфаузиид. Нерка явно специализируется на гипериидах, а чавыча и кижуч предпочитают кальмаров и рыб (Радченко, Рассадников 1997; Глебов, 2000). Таким образом, подобная ориентация в питания в значительной степени ослабляет конкуренцию за пищу.
Мы уже отмечали, что среди прочих видов нектона конкуренцию в питании кете могут составлять минтай, мезопелагические рыбы ^епоЬгасЫ^ пап-посЫг, S. leucopsarus и Leuroglossus schmidti) и кальмары. По данным Е.П.Дуле-повой (1998), суммарное суточное выедание эвфаузиид, амфипод, птеропод, копе-под и ойкоплевр этими представителями нектона составило в Беринговом море летом 1995 г. соответственно 1,36, 0,34, 0,75, 0,13 и 0,06 тыс. т. По нашим данным, выедание зоопланктона в верхней эпипелагиали юго-западной части моря минтаем, мезопелагическими рыбами и кальмарами летом 1993 г. оценивалось следующими цифрами: эвфаузиид — 1,89, амфипод — 0,47, птеропод — 1,04, копепод — 0,18 и ойкоплевр — 0,08 тыс. т. Таким образом, за время пребывания основной массы азиатской кеты старших возрастов в водах Берингова моря ее пищевыми конкурентами (кроме прочих лососей) в общем было потреблено летом 1993 г. 329,4 тыс. т, а летом 1995 г. — 237,51 тыс. т этих групп зоопланктона. Потребление планктонных и нектонных организмов тихоокеанскими лососями в эпипелагиали юго-западной части Берингова моря летом 1995 г. составило 226,5 тыс. т (Дулепова, 1998), а летом 1993 г. — 255,0 тыс. т. При этом, как было показано выше, выедание кетой тех же кормовых ресурсов летом 1993 и 1995 гг. составило более трети от этой суммы — соответственно 80,14 и 107,84 тыс. т. С учетом этого и приведенных выше данных суммарные оценки потребления основных пищевых объектов конкурентами кеты за 90-суточный интервал времени составят от 354,2 тыс. т (1995 г.) до 504,2 тыс. т (1993 г.).
Суммарная биомасса всех групп макропланктона в верхней эпипелагиали летом 1995 г. была более высокой по сравнению с предыдущим нечетным годом (в тихоокеанских водах летом 1995 г. — 19867 тыс. т против 14044 тыс. т в 1993 г. и в Беринговом море 25650 тыс. т против 21129 тыс. т). Однако, как отмечает В.П.Шунтов с соавторами (1997), величины биомассы планктонных организмов не всегда в полной мере отражают темп и степень использования кормовой базы потребителями, так как при этом не учитывается продукция планктона. По данным тех же авторов, средний Р/В-коэффициент зоопланктона в Беринговом море в летний период составляет 3,2-4,0, что соответствует высокому уровню продуцирования органической массы.
Исходя из приведенных значений можно сделать заключение, что в трофической структуре юго-западной части Берингова моря кета в период сезонного нагула занимает одно из ведущих мест. Конкурентные отношения за пищу между ней и наиболее массовыми представителями нектона вполне возможны на уровне таксономических групп зоопланктона — в первую очередь здесь следует отметить крылоногих моллюсков.
Осреднение данных для тихоокеанских вод восточной Камчатки позволяет представить схему трофических связей кеты в летний период в следующем виде: как и в Беринговом море, здесь основной пресс со стороны кеты приходится на птеропод. Менее значима роль эвфаузиид, молоди рыб и кальмаров. Напротив, потребление ойкоплевр более чем в 3 раза превосходит величину, полученную для юго-западной части Берингова моря (рис. 6). Среднесуточное потребление корма одной особью кеты было меньше, чем в Беринго-
 кетой кормовых организмов при среднем для первой половины 1990-х гг. уровне биомассы кеты")
Рис. 6. Трофические связи кеты в эпипелагиали тихоокеанских вод восточной Камчатки в период летнего нагула. Цифрами обозначено выедание (тыс. т) кетой кормовых организмов при среднем для первой половины 1990-х гг. уровне биомассы кеты
Fig. 6. Trophic linkages of chum salmon in epi pelagic layer of Eastern Kamchatka offshore waters during the summer period. Numbers designate consumption of prey
organisms (in thousands of metric tons) by chum salmon. An average biomass of chum salmon and its daily rations during the first half of 1990s were used
Таким образом, обобщая полученные оценки, можно заключить, что трофические связи кеты в юго-западной части Берингова моря и в водах восточной Камчатки базируются на крылоногих моллюсках, эвфаузиидах и в меньшей степени на гипериидах и представителях нектона. В целом группа лососей в период сезонного нагула оказывает заметное влияние на планктонные сообщества Берингова моря и тихоокеанских вод восточной Камчатки. Их пресс на планктон, по нашим оценкам, сопоставим с прессом остального эпипелагического нектона. При этом кета, самый массовый представитель тихоокеанских лососей в Беринговом море, среди этой группы рыб является в этом смысле лидером.
Пропорционально составу рационов кеты, а точнее — в зависимости от трофического уровня, с которого поступает энергия, ее можно отнести к третичным и одновременно к четвертичным консументам, т.е. к вторичным или третичным хищникам (Дулепова, 1998). Пластичность, проявляемая кетой в выборе объектов питания, позволяет в определенной степени избежать конкурентных отношений за пищу с другими представителями сообщества эпипелагиали.
Трофические связи кеты в нектонных сообществах южной части Охотского моря и тихоокеанских вод Курильских островов
В период преданадромных миграций кета в рассматриваемом районе продолжает активно питаться (Волков, 1996). Как мы отмечали выше, в это время питание созревающей кеты может значительно отличаться от питания неполовозрелых рыб. С другой стороны, заметны и региональные различия в питании как созревающих, так и неполовозрелых рыб по сравнению с питанием кеты в Беринговом море и тихоокеанских водах Камчатки. Наряду с характерными для Берингова моря и восточнокамчатских вод группами зоопланктона, такими как эвфаузииды, птероподы и гиперииды, в рационе кеты (главным образом половозрелой) отмечались в значительных количествах оболочники, медузы и саль
вом море, — 38 г ( за период летнего нагула — 3,42 кг).
пы. Спектры питания кеты с размерами тела заметно расширялись: если в рационах неполовозрелой кеты (размеры 31-40 и 41-50 см) встречались 3-4 группы пищевых организмов, то у половозрелых рыб, которые абсолютно преобладают среди кеты с размерами тела 51-60 и 61-70 см, число компонентов питания достигало 10-12.
Максимальное потребление кормовых ресурсов кетой в сахалино-куриль-ском регионе наблюдалось во время "подъема" хода осенней кеты — в августе—сентябре. Так, суточное выедание зоопланктона и нектона в 1994 г. в Охотском море составило 1,49 тыс. т, а в тихоокеанских водах — 2,88 тыс. т. В подавляющей массе (98 %) здесь присутствовали половозрелые рыбы. Так же как и в летний период 1993 и 1995 гг., именно половозрелая кета наиболее интенсивно питалась обводненными гидробионтами: потребление медуз за сутки в сумме по всей акватории съемки составило 0,9 тыс. т. Помимо этого, кета интенсивно выедала эвфаузиид, гипериид, птеропод и нектон. Причем в тихоокеанских водах молодь рыб и кальмаров по величине суточного потребления их кетой занимала лидирующую позицию — в сумме 0,73 тыс. т.
Величина выедания кетой кормовых ресурсов планктона и нектона за весь период исследований составила: в 1993 г. — 82,0 тыс. т, в 1994 — 261,9 и в 1995 г. — 208,9 тыс. т. Кроме того, в летний период 1993 г. в водах южной части Охотского моря в значительных количествах присутствовала посткатад-ромная молодь кеты. С учетом потребления ею за сутки 0,01 тыс. т зоопланктона суммарная оценка выедания кетой кормовых ресурсов в южной части Охотского моря летом 1993 г. составит 83 тыс. т.
Осреднение данных, полученных в первой половине 1990-х гг., позволяет представить схему трофических связей кеты в сахалино-курильском регионе в период ее летнего нагула (за который мы принимаем три летних месяца и первые две декады сентября) следующим образом. В это время основной пресс со стороны кеты был направлен на представителей зоопланктона — эвфаузиид, гипериид, птеропод и медуз (рис. 7). Из нектона кета предпочитала рыб головоногим моллюскам. Весьма существенным оказалось выедание сальп и оболочников (соответственно 7,6 и 5,9 тыс. т). Доля прочих групп планктонного сообщества, за исключением копепод и гребневиков, не превысила 0,2-0,5 % (0,3-0,8 тыс. т) от суммарной оценки сезонного рациона.
Суммарное потребление кетой корма за сезон составило 153,4 тыс. т, что соответствует 1,4 тыс. т в сутки. Для летнего периода первой половины 1990-х гг. суточное потребление корма одной особью кеты оценено нами в 54,6 г, что за период нагула составляет 6,1 кг корма.
Вторая половина 1990-х гг. была отмечена увеличением численности кеты (в первую очередь ее сеголеток), учитываемой в южной части Охотского моря и сопредельных тихоокеанских водах. Пропорционально возросло и потребление ею зоопланктона и нектона. Так, летом 1999 г. суточное выедание кетой кормовых ресурсов в этом регионе оценивалось в 0,51 тыс. т, что в пересчете только на период съемки составило 20,5 тыс. т.
В летний период конца 1990-х гг. основной пресс со стороны половозрелой кеты был направлен на нектон (выедание рыб за сутки составило 253,2 т), а из зоопланктона — на оболочников (148,7 т) и гипериид (52,5 т за сутки). Заметными в рационах кеты были эвфаузииды (в сумме для всех групп — 30,9 т).
Во время осенних исследований в суммарных рационах кеты абсолютно доминировали гиперииды (выедание 2,0-2,2 тыс. т в сутки). Основное воздействие на эту группу зоопланктона оказывали сеголетки кеты (выше мы уже отмечали, что именно гиперииды являются излюбленной пищей посткатадром-ной молоди кеты). Вторыми (третьими) по значимости в питании сеголеток кеты в этом регионе, как и в предыдущий период исследований (первая половина 1990-х гг.), были копеподы (0,65-1,40 тыс. т) и хетогнаты (0,54-0,61 тыс. т).
Рис. 7. Трофические связи кеты в эпипелагиали сахалино-курильского региона в период сезонного нагула. Цифрами обозначено выедание (тыс. т) кетой кормовых организмов при среднем для первой половины 1990-х гг. уровне биомассы кеты
Fig. 7. Trophic linkages of chum salmon in epi pelagic layer of the Sakhalin-Kuril region in the during the first half of 1990s. Numbers designate consumption of prey organisms (in thousands of metric tons) for average biomass by chum salmon in first half 1990s
В питании крупноразмерной кеты в первую очередь отметим нектон (0,230,51 тыс. т) и птеропод (0,14-0,16 тыс. т).
Суммарные оценки потребления зоопланктонных и нектонных организмов кетой в водах сахалино-курильского региона в период сезонного нагула позволяют представить схему трофических связей кеты в этом регионе для второй половины 1990-х гг. следующим образом. В целом за период нагула кета потребляет 400 тыс. т пищи в южной части Охотского моря и тихоокеанских водах Курильских островов. Основной пресс с ее стороны приходится на гипериид, копепод, эвфаузиид и хетогнат (рис. 8). Первые две группы зоопланктона наиболее интенсивно выедаются сеголетками кеты. Оценки выедания хетогнат (40,2 тыс. т), птеропод (22,2 тыс. т), нектона (рыб и кальмаров в сумме 32,5 тыс. т) и оболочников (5,2 тыс. т) практически тотально соответствуют уровню потребления их крупноразмерной, главным образом осенней созревающей, кетой. То же справедливо для ойкоплевр (12,2 тыс. т) и гребневиков (17,8 тыс. т).
Сопоставление суммарных оценок, полученных для первой и второй половины 1990-х гг., показывает, что уровни потребления нектона и птеропод кетой абсолютно совпадают. В противоположность этому для конца 1990-х гг. отмечен существенный рост потребления гипериид и копепод (что в первую очередь мы связываем с ростом массы сеголеток в регионе), при прежнем соотношении в рационах кеты эвфаузиид. К концу десятилетия также увеличилось потребление (практически исключительно половозрелой кетой) хетогнат, ойкоплевр и гребневиков при снижении (с 26 до 1 тыс. т) выедания медуз.
Итоговые оценки выедания зоопланктона и нектона кетой в сахалино-ку-рильском регионе второй половины 1990-х гг. превысили уровень начала десятилетия в 2,5 раза.
Рис. 8. Трофические связи кеты в эпипелагиали сахалино-курильского региона в период сезонного нагула. Цифрами обозначено выедание (тыс. т) кетой кормовых организмов при среднем для второй половины 1990-х гг. уровне биомассы кеты
Fig. 8. Trophic linkages of chum salmon in epi pelagic layer of southern part of Okhotsk Sea and Pacific waters of Kuril Islands during the second half of 1990s. Numbers designate consumption of prey organisms (in thousands of metric tons) by chum salmon. An average biomass of chum salmon and its daily rations during the second half of 1990s were used
Обобщая полученные данные по сахалино-курильскому региону, выедание кормовых ресурсов кетой за время ее пребывания здесь можно оценить в 300350 тыс. т в первой половине 1990-х гг., а во второй половине 1990-х гг. — не менее 400-500 тыс. т.
Для северной части Охотского моря более полная информация получена во второй половине 1990-х гг. (1997-2000). В то же время материалы по питанию кеты собирались в северо-восточной части моря и в предыдущие годы (1991, 1994-1996). Так, суммарные цифры суточного выедания пищи кетой в северовосточной части моря летом 1995 г. абсолютно совпали с результатами, полученными в этом же году для его южной части и тихоокеанских вод Курильских островов (0,90 и 0,91 тыс. т при биомассах кеты соответственно 23,7 и 24,4 тыс. т). Этот факт, несомненно, свидетельствует в пользу большой значимости для летнего нагула кеты сравнительно необширного по площади (по отношению к суммарной площади районов южной части моря и сопредельных тихоокеанских вод) западнокамчатского района.
Как отмечено выше, нектонная составляющая в питании кеты в северной части Охотского моря более значительна, чем в южной. По данным летних исследований 1997 г., потребление молоди рыб (главным образом сельди и минтая) в североохотских районах кетой размерной группы 51-60 см в сумме составило 0,443 тыс. т и 0,726 тыс. т рыбами, размеры которых превышали 60 см.
Сеголетки кеты летом 1997 г. питалась исключительно молодью минтая, сельди и песчанки. Однако ввиду незначительной биомассы посткатадромной молоди кеты выедание ею нектона составило лишь 3,6 т за сутки. Схожие оценки потребления рыб (3,9 т) получены и для кеты размерной группы 31-40 см, которая была представлена главным образом рыбами второго морского года жизни.
Осенью 1998-1999 гг. основными потребителями зоопланктона и нектона в северной части Охотского моря были сеголетки (размерная группа 21-30 см) и половозрелая кета (рыбы с размерами тела более 61 см). Суточное потребление корма кетой в северной части Охотского моря составило 0,51 и 0,15 тыс. т соответственно в 1998 и 1999 гг. При этом осенью 1998 г. 60 % этой оценки пришлось на сеголеток, а в 1999 г. основной пресс (90 %) на зоопланктон и нектон оказывали рыбы старших возрастов.
Обобщение оценок выедания пищи кетой в северной части Охотского моря в летний и осенний периоды второй половины 1990-х гг. позволило получить следующие результаты. В отличие от южной части моря, на первом месте по важности в питании кеты оказались рыбы (точнее молодь минтая, сельди, песчанки и в меньшей степени мойва и серебрянка) — 105 тыс. т (рис. 9).
 кетой кормовых организмов при среднем для второй половины 1990-х гг. уровне биомассы кеты")
Рис. 9. Трофические связи кеты в эпипелагиали северной части Охотского моря в период сезонного нагула. Цифрами обозначено выедание (тыс. т) кетой кормовых организмов при среднем для второй половины 1990-х гг. уровне биомассы кеты
Fig. 9. Trophic linkages of chum salmon in epi pelagic layer of northern part of Okhotsk Sea during the second half of 1990s. Numbers designate consumption of prey organisms (in thousands of metric tons) by chum salmon. An average biomass of chum salmon and its daily rations during the second half of 1990s were used
Относительно высокий уровень потребления эвфаузиид (22 тыс. т) и гипе-риид (14 тыс. т) практически абсолютно можно отнести на счет сеголеток кеты. Напротив, потребление нектона (включая 4 тыс. т кальмаров) было в основном связано с хищничеством кеты старших возрастов.
Еще одна особенность трофических связей кеты в северной части Охотского моря состояла в том, что выедание гребневиков, оболочников, хетогнат и медуз здесь оказалось несущественным по сравнению с южной частью моря: в суммарном сезонном рационе кеты лишь оболочники (главным образом Beroe cucumis) достигли уровня 0,6 тыс. т, при несравненно меньших оценках потребления других групп желетелых гидробионтов (рис. 9).
Причина различий рационов половозрелой кеты на севере и юге моря, на наш взгляд, заключается в особенностях питания осенней кеты, которая осенью по биомассе абсолютно доминирует в южной части Охотского моря и сопредельных водах Тихого океана и в то же время относительно малочисленна на севере моря, где нагуливаются главным образом летняя кета и сеголетки.
Потребление зоопланктона и нектона кетой в летний (июль—август) и осенний (сентябрь—октябрь) период второй половины 1990-х гг. составило 190 тыс. т, что соответствует выеданию 1,6 тыс. т корма в сутки. Заметим, что эта оценка не охватывает начало летнего периода и конец осеннего, так как в это время исследования в северной части моря проводились лишь в отдельных районах. Тем не менее учитывая, что к началу лета (конец мая — июнь) максимум биомассы кеты наблюдается в западнокамчатских водах, а к концу осени (ноябрь) — в центральной котловине, и используя осредненные данные по биомассам и суточным рационам кеты в указанных районах, к суммарной цифре потребления пищи можно с уверенностью добавить еще 30-50 тыс. т. В зимний период кета покидает воды северной части моря и соответственно не использует кормовые ресурсы этого региона.
Таким образом, в конце 1990-х гг. за период сезонного нагула в северной части Охотского моря кета потребляла более 200 тыс. т корма. Половина этой величины приходилась на нектон (а именно на молодь рыб), четвертая часть — на крылоногих моллюсков и еще четверть — на прочих представителей зоопланктона.
Обобщая полученные данные для южной и северной части Охотского моря, можно, с одной стороны, констатировать большее разнообразие трофических связей кеты по отношению к таковым в Беринговом море, а с другой стороны, отметить региональные особенности ее питания. Так, потребление желетелых организмов характерно (здесь мы имеем в виду не единичный, а массовый характер питания отмеченными объектами) только для созревающей кеты в южной части моря и сопредельных вод Курильских островов и не отмечается в других регионах. В северной части Охотского моря для кеты свойственна "не-ктонная" (точнее " нектонно-птероподная") направленность питания, а не " птеро-подная", как в западной части Берингова моря.
Суммарные оценки потребления кетой ресурсов зоопланктона и нектона в сахалино-курильском регионе для летне-осеннего периода первой половины 1990-х гг. соответствуют уровню выедания кетой корма в камчатско-берингово-морском регионе в это же время (соответственно 153,4 и 159,6 тыс. т). Во второй половине 1990-х гг. отмечено существенное увеличение биомассы кеты в Охотском море, что отразилось и на уровне потребления ею кормовых ресурсов: только в южной части моря и сопредельных водах оценки выедания корма кетой в 2,5-3,0 раза превосходят цифры, полученные для первой половины 1990-х гг. В северной части моря потребление зоопланктона и нектона к концу 1990-х гг. составило 200-250 тыс. т, что, очевидно, так же как и для южной части моря, выше уровня первой половины десятилетия.
Для оценки роли кеты как потребителя кормовых ресурсов верхней эпипе-лагиали Охотского моря приведем количественные оценки выедания зоопланктона и нектона ее конкурентами. В конце 1980-х гг. в ихтиоценах эпипелагиали Охотского моря в период сезонного нагула лососей на юге абсолютно доминировала дальневосточная сардина иваси, а на севере моря — минтай. При сравнительно невысокой интенсивности питания (12-47 %о о) 20-30 % рациона сардины иваси составляет фитопланктон, а остальную часть — мелкие планктонные ракообразные, длина которых в основном не превышает 3 мм (Лапко, 1996). Таким образом, конкурентные отношения за пищу между кетой и сардиной иваси могут не учитываться как существенные. Главная роль в трофическом процессе принадлежит минтаю, как наиболее активному мигранту и основному переносчику ихтиомассы в системах шельфа и открытых вод. Несмотря на существенное сокращение численности, этот вид по-прежнему является основным потребителем кормовых ресурсов в Охотском море. Помимо него, в группу лидеров по потреблению зоопланктона и в меньшей степени нектона следует включить серебрянку, сельдь, мойву и лососей.
Выедание кормовых объектов в летний период второй половины 1980-х гг. этими потребителями составило 46,6 млн т (Лапко, 1996). Из них на долю минтая пришлось 79,0 % (36,9 млн т), сельди — 7,5 % (3,5 млн т), серебрянки — 12,1 % (5,6 млн т), мойвы и лососей — по 0,6 % (0,26 и 0,28 млн т). В этой суммарной оценке на долю горбуши пришлось 0,5 % (0,22 млн т), а на долю кеты — почти 0,1 % (0,04 млн т). Судя по этим цифрам, среди указанных видов кета играла весьма скромную роль как потребитель корма.
Совсем иные соотношения были получены спустя десятилетие — в летний период конца 1990-х гг. Вследствие значительного уменьшения численности (биомассы) минтая, снизилось и потребление им корма — до 12,6 млн т (39,1 % суммарной оценки выедания в 32,2 млн т). Лидировать по потреблению корма стала сельдь — 14,6 млн т, или 45,4 %. В пять раз сократилась величина потребления корма серебрянкой — 2,3 млн т (7,1 %) — при более чем двухкратном росте этой величины у мойвы и группы лососей — соответственно 1,4 млн т (4,4 %) и 1,3 млн т (4,0 %). Из последней оценки на долю горбуши пришлось 0,7 млн т (2,2 %), а на долю кеты — 0,5 млн т (1,6 %).
Таким образом, спустя десятилетие значимость обоих видов лососей (горбуши и кеты) в суммарном летнем потреблении зоопланктона и нектона в Охотском море возросла по сравнению с периодом второй половины 1980-х гг. более чем на порядок.
А.Н.Старовойтов