
ФОРМИРОВАНИЕ ЖИЗНЕННОЙ СТРАТЕГИИ В ПОПУЛЯЦИИ СИМЫ РЕКИ КОЛЬ (ЗАПАДНАЯ КАМЧАТКА)
Выбор жизненной стратегии по анадромному или резидентному типу
у лососевых рыб определяется в период пресноводной фазы жизненного
цикла, связан с метаболическими процессами и перестройкой осморегу-
ляторной системы, причем среда действует как переключатель программы
индивидуального развития (Павлов, Савваитова, 2010 и др.).
Изученность особенностей дифференцировки у разных видов проходных лососевых рыб
неодинакова, поэтому неясно, насколько универсальны механизмы форми-
рования жизненной стратегии. Из тихоокеанских лососей сима наиболее
связана с пресными водами – помимо проходной формы, известны озерная
и жилая речная формы, представленные самцами и самками (GGrroooott,,, MMMaaarrr--Margolis,
1991). Все разнообразие форм приурочено к южным участкам ареала
вида; на севере ареала, на Камчатке, у симы обнаружены проходные сам-
цы и самки и карликовые самцы. В 2003–2008 гг. исследована популяция
симы р. Коль: проходные производители и разновозрастная молодь, про-
веден сравнительный анализ разных группировок (пестряток, покатников-
смолтов и карликовых самцов) по размерно-весовым характеристикам,
возрасту, темпу роста, строению чешуи, упитанности, состоянию половых
желез.
Проходная сима р. Коль представлена рыбами в возрасте 1.1+, 1.2+, 2.1+
и 2.2+, карликовые самцы – 1+, 2+ и 3+, неполовозрелая молодь (пестрят-
ки и смолты) – 0+, 1+ и 2+. Учитывая длительность пресноводной фазы
жизненного цикла проходных рыб (не более 2 лет) и предельный возраст
карликовых самцов (3+), можно говорить, что процессы дифференциации
в поколении происходят в течение первых двух лет жизни.
Разнокачественность по длине тела и темпу роста заметна у сеголеток (0+) уже к концу первого лета жизни. В начале осени у особей с наиболь-
Проблемы сохранения и функционирования особо охраняемых природных территорий 223
шим темпом роста и размерами тела проявляются признаки смолтифика-
ции – именно они следующей весной превратятся в смолтов и скатятся в море (рис.). У будущих карликовых самцов темп роста ниже и размеры меньше, чем у будущих покатников-смолтов. Осенью первого лета жизни будущие карликовые самцы имели гонады на II-III стадии зрелости, тогда как гонады самцов пестряток и будущих смолтов находились исключи-
тельно на II стадии зрелости, их половые клетки были представлены спер-
матогониями первых порядков. Яичники всех самок в этом возрасте были на II стадии зрелости первой ступени цитоплазматического роста ооцитов. Во второе лето жизни пестряток (возраст 1+) обнаружена сходная картина дифференцировки – наиболее крупные, быстрорастущие особи являются будущими смолтами, самцы с промежуточными показателями темпа ро-
ста превратятся на следующий год в карликовых самцов. Состояние го-
над в группировках в 1+, в целом, сходно с таковым в 0+. В то же время в конце второго лета жизни среди наиболее медленно растущих пестряток остаются только самцы. Поэтому на следующий год, когда рыбы достиг-
нут возраста 2+, после ската в море смолтов, в реке останутся только сам-
цы – одна часть из них половозрелые карлики, другая часть – с гонадами на II стадии зрелости. Неполовозрелые самцы в возрасте 2+ в дальнейшем так и останутся в реке, став половозрелыми на следующий год, в возрасте 3+ (рис.).
В р. Коль карликовые самцы симы принимают участие в нересте с крупными проходными самками – на нерестилищах в период нереста на 1 проходную самку приходится 1 проходной самец и 7–15 разново-
зрастных карликовых самцов. Сима в р. Коль нерестится в узких горных ручьях, порой самка строит нерестовый бугор в узких участках ручьев шириной менее 1 м, поэтому мелкие карликовые самцы могут играть основную роль в оплодотворении икры. После окончания нереста про-
ходные самцы и самки быстро погибают, но карликовые самцы еще долго остаются на нерестилищах и даже скатываются из нерестовых ручьев в основное русло реки. В семенниках карликовых самцов после нереста (стадия VI-II) обнаружены единичные делящиеся сперматогонии, а в ка-
-) нальцах – невыметанные и фагоцитируемые спермии. Гистологический анализ гонад показал, что предположительно у этих рыб возможно по-
вторное созревание, однако за все годы исследований в р. Коль повторно-
го нереста карликовых самцов не обнаружено. Возможность выживания карликовых самцов после нереста и повторный нерест отмечались для рек бассейна Японского моря (Иванков и др., 1977) и Сахалина (Крыхтин, 1962), однако на Камчатке, в северной части ареала вида, истощенные после нереста карликовые самцы, по всей видимости, не могут перенести продолжительной суровой зимы. Обнаруженные в популяции симы из р. Коль закономерности формиро-
вания анадромного и резидентного образа жизни сходны с таковыми у ат-
лантического лосося (Thorpe, 1987) и кижуча (Павлов и др., 2007). Эти виды
строго анадромные (созревание самок в пресных водах при возможности
выхода в море является редким исключением), для них характерно, что со-
матический рост, требующий значительной энергии, у наиболее быстрора-
стущей в поколении молоди существенно замедляет гаметогенез, гонады
самцов и самок остаются на II стадии развития. Быстрый рост стимулирует
запуск процесса смолтификации и предопределяет выбор анадромного типа
жизненной стратегии. Известно, что у многих видов лососевых рыб смол-
тификация возможна лишь при достижении определенных «минимальных
критических размеров» (Черницкий, 1993), поэтому именно быстрорасту-
щая молодь, в первую очередь, становится в дальнейшем проходной частью
популяции. У самцов с несколько замедленным темпом роста, которые не
достигли размеров для смолтификации, начинается активный гаметогенез,
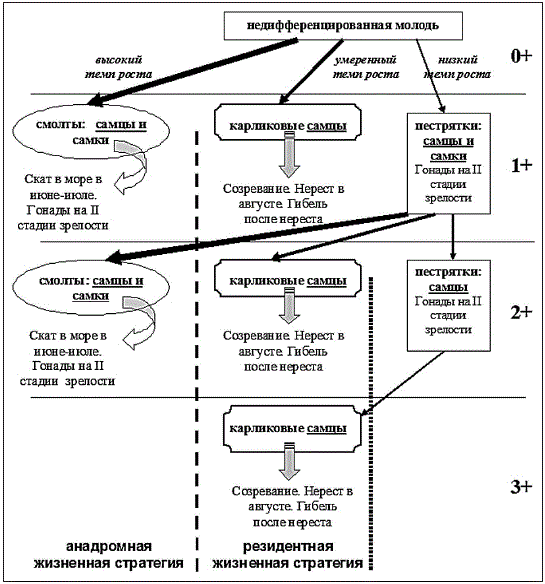
Схема дифференциации в поколении симы р. Коль
Проблемы сохранения и функционирования особо охраняемых природных территорий 225
приводящий, в конечном счете, к созреванию в пресной воде и реализации
резидентного типа жизненной стратегии.
Таким образом, важнейшими факторами, определяющими дифференци-
ацию молоди симы и направление онтогенетического развития, являются
темпы роста и гаметогенеза. Процессы внутрипопуляционной дифференци-
ровки в значительной степени определяются условиями обитания в ранний
пресноводный период, главным образом, трофическим фактором (Павлов
и др., 2007). Экосистемы лососевых рек Камчатки характеризуются высо-
кой продуктивностью за счет огромной численности нерестящихся в них
стад тихоокеанских лососей и привнесения ими биогенов морского проис-
хождения. В то же время из-за межгодовых колебаний численности тихоо-
кеанских лососей и абиотических факторов внешней среды наблюдается
колебательная природа формирования жизненной стратегии в популяции
симы: как правило, в четные, урожайные годы, рост молоди симы выше,
большая часть поколения уходит в море или превращается в карликовых
самцов в возрасте 1+, в нечетные годы возрастает доля старшевозрастных
смолтов и карликовых самцов в возрасте 2+ и 3+.
М.А. Груздева*, А.М. Малютина*, К.В. Кузищин*, Н.В. Белова*, С.В. Пьянова**
*Московский государственный университет им. М.В. Ломоносова (МГУ)
**Всероссийский научно-исследовательский институт рыбного хозяй-
ства и океанографии (ВНИРО), Москва