
Возможности раскрытия ростовой потенции у радужной форели в УЗВ и открытых рыбоводных системах
Радужная форель, как любой представитель сообщества гидробионтов, являясь пойкилотсрмным животным, испытывает воздействие множества абиотических и биотических факторов. Среди основных принято учитывать (по классификации У. Хоара и соавторов, 1983) температуру воды как основной направляющий развитие организма фактор. Кислородный и соленостный режим рассматриваются как основные лимитирующие факторы.
Все остальные абиотические факторы отнесены к сопутствующим. Среди биотических факторов основным признается режим кормления, который в аквакультуре следует рассматривать в разрезе применяемой системы нормирования. Нормирование кормления, как известно, включает комплекс элементов: состав рецептуры корма, суточная доза корма, кратность кормления, размер кормовых частиц, метод и способ кормления [1]. Остальные биотические факторы, такие, как плотность посадки, половой и возрастной состав рыб, моно- и поликультура, следует признать сопутствующими в процессе усвоения корма.
Взаимодействие всех перечисленных факторов раскрывается в качественной и количественной сторонах двух основных по рыбохозяйственной значимости процессов: роста и созревания. Причем, как показала теория и практика аквакультуры, равновесного соотношения этих процессов по величине отвлекаемой энергии общего обмена в течение жизненного цикла рыб не отмечается.
У ювенильных рыб наибольшее количество энергии общего обмена направляется на рост. У половозрелых рыб большая часть энергии направляется на рост только в период, когда отмечается протоплазматический рост половых клеток. При наступлении периода трофоплазматического роста половых клеток структура обмена перестраивается в направлении генеративного. Поэтому в практике рыбоводства для достижения ростового эффекта ориентируются на те периоды или циклы выращивания рыб, когда они представлены группами неполовозрелых рыб или же учитывается определенная часть периода посленерестового (межнерестового) нагула. Исследованиями установлено, что скорость роста неполовозрелых и половозрелых рыб может быть сопоставима по величине в отмеченные выше периоды.
Анализируя или управляя процессом выращивания рыбы, необходимо опираться на основные теоретические положения, подтверждаемые практикой аквакультуры. Во-первых, на правило Вант-Гоффа-Аррениуса: каждое последующее повышение температуры воды на 10 °С в сторону оптимального для рыб диапазона сопровождается увеличением предельной скорости роста в 2-3 раза [2]. К примеру, если мы признаем, что некоторый рост радужной форели проявляется при температуре 2-3 °С, то следует ожидать его увеличения в 2-3 раза, когда температура воды повысится до 12-13 °С. Учитывая, что оптимальная температура воды для форели 14-18 °С, последующее увеличение скорости роста может быть 1,2-2-кратным. Несмотря на то что в нормативной литературе зафиксировано, что зимовка форели в рыбоводных хозяйствах, где температура воды в зимний период опускается ниже 1 °С, сопровождается увеличением массы рыб на 30 %, следует учитывать, что этот прирост фактически приходится на переходные периоды понижения температуры воды ниже 5 °С осенью и повышение в сторону 5 °С весной. Именно при такой температуре проводятся, как правило, последняя осенняя и первая весенняя сортировки и контрольное взвешивание [2].
В тех же источниках указано, что если зимой температура воды 3-5 °С, то прирост за период должен составлять 100 %. Если скорость роста фиксируют при такой температуре, то следует ожидать, что увеличение ее в 2-3 раза придется на
период оптимальной температуры воды. Это может означать, что если при температуре воды 3-5 °С, отмечаемой в течение 4 мес, произошло удвоение массы форели, то в перпод оптимальной температуры удвоения или даже утроения массы рыб следует ожидать в течение 1,5-2 мес. Поэтому если в течение вегетационного сезона (от 5 °С весной до 5 °С осенью) продолжительностью, например, 6 мес. перпод с температурой 14-18 °С составит 4 мес., то можно рассчитывать на возрастание массы рыб минимум в 4 раза в самый благоприятный период. На осенние и весенние периоды с переходной температурой воды следует планировать также определенный прирост массы рыб. Но весной он будет по отношению к стартовой массе рыб, осенью к массе рыб, достигнутой к концу периода интенсивного роста. Тем не менее, можно приближенной величиной оценивать в данном примере скорость увеличения массы форели в 5-6 раз. Если обратиться к литературным, в том числе нормативным, источникам, то можно принять за близкую к достоверной указанную скорость роста: прп посадке весной на выращивание в пруды, садки, бассейны годовиков форели массой 25-30 г масса товарной порцпонной форели составляет 125-150 г, прп посадке весной на нагул годовиков форели массой 80-100 г к осени рыбы достигают средней массы 400-600 г [3]. При этом следует отметить, что на практике трудно выделить четкие границы периодов переходной п стабильной оптимальной температуры.
Во-вторых, следует опираться на закон Ю. Лпбпха, который учитывает совокупное действие всех экологических факторов п проявляемое в величине экологического коэффициента роста - Кэ [2]. Однако это действие надо рассматривать по двум позициям.
Первая позицпя согласно закону Ю. Либиха рассматривается как способ абсолютного лимитирования. В этом случае общее продуктивное действие всех факторов определяется только одним, имеющим минимальное значение. Формулу экологического коэффициента роста следует рассматривать как произведение множества частных экологических коэффициентов:
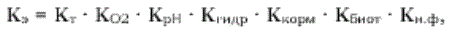
где Кт - коэффициент, определяющий степень влияния температуры воды на рост; К02 - коэффициент, определяющий степень влияния концентрации кислорода в воде на рост; Крн - коэффициент, определяющий степень влияния активной реакции среды на рост; КГиДР - коэффициент, определяющий степень влияния химических элементов на рост; Ккорм - коэффициент, определяющий степень влияния кормления на рост; Кбиот - коэффициент, определяющий степень влияния уровня биотехники на рост; Кн.ф - коэффициент, определяющий степень влияния неучтенных факторов на рост.
Тогда можно предположить, что прп максимальной величине всех факторов, кроме одного, результат будет следующим:
Кэ = 1 • 1 • 1 • 1 • 1 • 1 • 0,7 = 0,7.
Так закон работает, если в качестве объекта рыба, а форель таковой является, прошедшая длительную доместикацию, селекционные достижения очевидны, технические параметры рыбоводных систем устоявшиеся, биотехника на стабильном уровне.
В том случае если указанные параметры нестабильны, тем более, если объект выращивания на первых этапах доместикации, то следует принимать другой способ, учитывающий совокупное действие лимитирующих факторов. Это вторая сторона закона Ю. Либиха. В этом случае величина экологического коэффициента роста рассчитывается путем перемножения всех частных коэффициентов.
Например, Кэ = 1 • 1 • 1 • 1 • 0,8 • 0,7 • 0,8 = 0,448. Если мы применим в этой формуле подход по первому способу, то величина Кэ была бы 0,7.
Основываясь на вышеприведенном материале, можно прогнозировать рост рыб, анализировать результаты, устанавливать причины, определившие их.
Расчетным механизмом, позволяющим конкретизировать результаты и прогноз, является формула общепродукционного коэффициента массонакопления [2]. Базовый вид формулы:
Км = Кг*'Кэ, (1)
где Кг - генетический коэффициент роста; Кэ - экологический коэффициент роста.
Если подход к расчету величины Кэ приведен ранее, то относительно величины Кг можно обратиться к источнику [2].
Для радужной форели в одних вариантах приводится значение (0,078), близкое к семге (0,074), в других фигурирует значение Кг для полупроходной формы радужной форели - стальноголового лосося, составляющее 0,102. Опираясь на данные еще одного источника, можно согласиться, что величина Кг для радужной форели в интервале 0,10-0,11 [4].
Таким образом, при величине Кг = 0,1 и величине Кэ = 1 (идеальные условия для отселектированного материала) величина Км = 0,1.
Далее, если воспользоваться расчетным вариантом формулы (2):
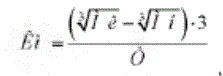 , (2)
, (2)
где Км - общепродукционный коэффициент массонакопления; Мн и М к — масса рыб начальная и конечная, г; Т - период выращивания, сут.
Можно преобразовать ее в прогнозный вариант, представленный формулой (3):
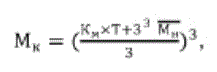 (3)
(3)
Далее нами предлагается к рассмотрению матрица, учитывающая возможное действие каждого частного экологического фактора (табл. 1).
Таблица 1
Возможные значения частных экологических факторов и экологического коэффициента роста, рассчитанного но способу абсолютного лимитирования
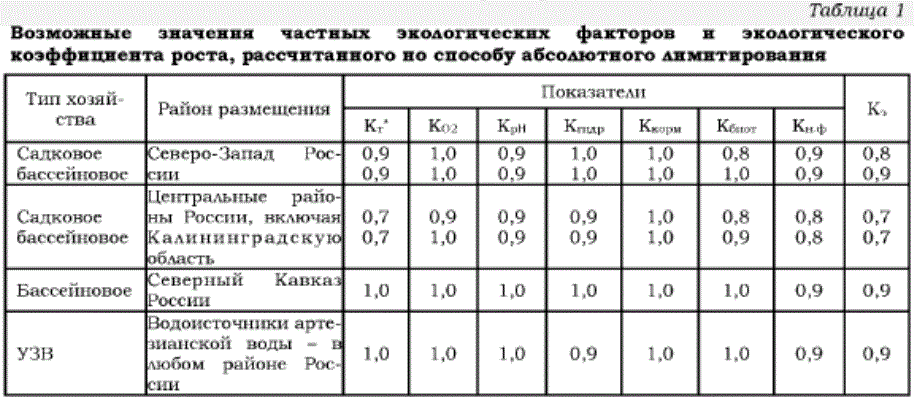
Основываясь на данных табл. 1 и оперируя формулами (1) и (2), можно рассчитать прогнозируемую массу посадочного материала и товарной рыбы. В качестве исходного посадочного материала учитываем 1 г мальков, адаптационная система которых развита достаточно, чтобы адекватно реагировать на изменения в окружающей среде. Расчетные результаты представлены в табл. 2.
Таблица 2
Ожидаемая средняя масса форели в разных тинах рыбоводных хозяйств, размещаемых на территории России, г
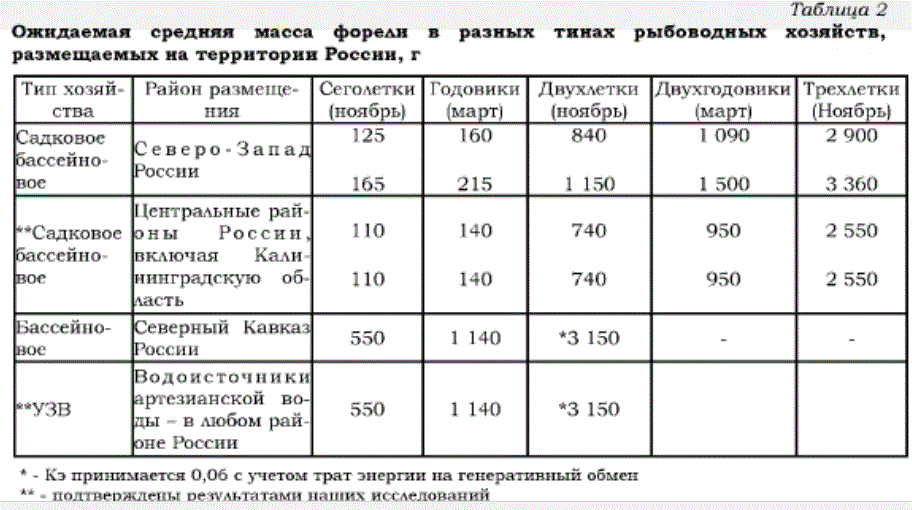
Для районов Северо-Запада продолжительность периода с благоприятной для роста температурой воды составляет 150 сут, в центральной части России 165 сут, а на Северном Кавказе 240 сут, в рамках указанных в табл. 2 временных границ. Но при этом учитывается, что водоисточники, снабжающие хозяйства водой, имеют среднегодовую температуру воды около 14 °С. В УЗВ круглый год устанавливается благоприятный для роста температурный режим.
Приведенные расчетные данные отражают ростовую потенцию радужной форели и могут закладываться при разработке технологической части проектов строительства рыбоводных хозяйств. Возможные отклонения в конечных результатах рыбоводного процесса непосредственно в период эксплуатации являются результатом отклонения в величине одного или нескольких частных экологических факторов.
Аспирант K.A. Молчанова, профессор Е.И. Хрусталев, доцент Т.М. Курапова (Калининградский государственный технический университет») кафедра аквакультуры, тел. 8 (4012)-99-53-53 E-mail: ksenia.elfimovatogmail.com