
Влияние комбикормов различного состава на ростовые процессы радужной форели
Решение вопросов рационального использования ресурсов внутренних водоемов относится к числу важнейших актуальных направлений современной биологии, включающих исследования в области ихтиологии, гидробиологии, физиологии, биохимии. Снижение вылова ценных видов рыб из естественных водоемов компенсируется их интенсивным выращиванием в искусственных условиях, в большей степени в морских акваториях [Emre et al., 2007; FAO, 2009]. В то же время наличие большого количества глубоководных озер с чистой водой на северо-западе России позволяет развивать садковое рыбоводство радужной форели в открытых пресных водоемах.
Для реализации основной задачи форелевых хозяйств, связанной с получением товарной продукции за максимально короткий период времени, в качестве источника пищи используют искусственные корма. Численность садковых хозяйств в последнее десятилетие резко возросла, в связи с чем увеличилась потребность в кормах, что привело к резкому дефициту сырья для их производства [FAO, 2008; Hua, Bureau, 2009]. Комбикорма для аквакуль-туры лососевых рыб производят преимущественно из отходов промыслового рыболовства. Экономически обоснованным альтернативным источником сырья служат продукты растительного происхождения (масла, протеин гороха, глютелин кукурузы и другие), которые, однако, не соответствуют кормовой базе хищных рыб из природных водоемов.
Состав пищи в первую очередь влияет на метаболизм рыб, который определяет интенсивность их роста и развития, а также качество реализуемой форелеводами продукции [Ruyter et al., 2010; Yun et al., 2011]. В настоящее время активно изучается влияние различных составов
комбикормов на морфометрические и физио-лого-биохимические характеристики культивируемых рыб, особенно лососевых, однако эти вопросы остаются все еще слабоизучен-ными [Tocher, 2003; Brown et al., 2010]. Целью настоящей работы было определение влияния комбикормов разного состава на темп роста радужной форели Parasalmo mykiss (Walbaum, 1792).
Исследование проводилось на базе непол-носистемного форелевого хозяйства, расположенного в северной части Ладожского озера (61°42'15" с.ш., 31°0'27" в.д.) (Республика Карелия, Россия). Изучены три группы неполовозрелых самок радужной форели Parasalmo mykiss (Walbaum, 1792) в возрасте 1+, не различающихся морфо-генетическими особенностями и культивированных в одинаковых условиях на разных комбикормах. В эксперименте применялись три вида коммерческих комбикормов (с одинаковыми кормовыми коэффициентами), наиболее часто используемые форелеводами Северо-Запада России. Рыб из одного садка, питающихся комбикормом № 1, в марте разделили на две группы (№ 1 и № 2) и культивировали их на комбикормах № 1 и № 2 соответственно. В конце июня форель группы № 2 случайным образом разделили на две группы (№ 2 и № 3), одну из которых - группу № 2 - продолжили кормить комбикормом № 2, а вторую группу -№ 3 - перевели на корм другого производителя (комбикорм № 3). Рыб выращивали в близкорасположенных садках с целью нивелирования неконтролируемых факторов. В третьей декаде каждого месяца проводили промеры групп рыб (по 50 особей в каждой группе) и фиксировали пробы кормов на биохимический анализ. В связи с длительным периодом исследования дважды провели смену фракции кормов, при этом марка комбикорма осталась прежней. С марта по июнь форель кормили кормами фракции А, размер гранул составлял 3 мм в диаметре; 1 июля произошла замена корма на фракцию В (4,5 мм); после 1 сентября рыб кормили комбикормами фракции С (6 мм).
Оценку длины, массы, прироста массы и длины радужной форели, наполненности кишечника, смертности рыб проводили по методике И. Ф. Правдина [1966] с учетом рекомендаций М. В. Мины с соавт. [Мина, Клевезаль, 1976], Л. П. Рыжкова и др. [2000]. В комбикормах определяли общий белок, липиды (структурные и запасные) и жирные кислоты общих липидов. Концентрацию белка оценивали по содержанию азота в пробе, полученного путем сжигания навески корма в колбе Кьельда-ля [Филиппович и др., 1975]. Для анализа ли-пидного состава кормов пробы фиксировали смесью Фолча (хлороформ : метанол в соотношении 2 : 1 по объему). Разделение общих липидов проводили методом тонкослойной хроматографии восходящим способом в системе растворителей: петролейный эфир : ди-этиловый эфир : уксусная кислота (в соотношении 90 : 10 : 1 по объему) при комнатной температуре [Шталь, 1965]. Концентрацию исследуемых липидных параметров определяли спектрофотометрическими методами [Сидоров и др., 1972; Епде1ЬгесМ et а1., 1974]. Фракционирование фосфолипидов осуществляли с помощью высокоэффективной жидкостной хроматографии. Подвижная фаза (элюент) состояла из смеси растворителей: ацетонит-рил : гексан : метанол : фосфорная кислота (918 : 30 : 30 : 17,5 по объему). Обработка хро-матограмм проводилась с помощью компьютерной программы «Мультихром-Аналитик, V. 1.5». Для анализа жирнокислотного состава комбикормов выделенные липиды подвергали прямому метилированию [Цыганов, 1971]. Полученные метиловые эфиры жирных кислот разделяли на хроматографе «Кристалл 5000» («Хроматек», Йошкар-Ола). Идентификацию жирных кислот проводили сопоставлением времени выхода пиков экспериментального образца и метчиков, а также путем расчета эквивалента длины цепи и сравнением его с табличными данными [Jamieson, 1975], количественный анализ осуществляли при помощи компьютерной программы «Программа подсчета хроматограмм». Полученные данные обрабатывали общепринятыми статистическими методами [Елисеева, 2007]. Исследования выполнены на научном оборудовании ЦКП НО Института биологии КарНЦ РАН.
Результаты и обсуждение
Интенсивность ростовых процессов радужной форели во многом определяется составом кормов, которые используются при ее выращивании. Для активного роста и развития культивируемых рыб необходим высокий уровень белка в корме, который должен использоваться именно для пластического обмена, а не для энергозатрат организма. Включение в корм соответствующих небелковых источников энергии, таких как липиды, позволяет оптимизировать химический состав кормов, поскольку липиды помимо энергетической выполняют в организме рыб ряд других жизненно важных функций: структурообразующую, регуляторную и прочие, к тому же они служат предшественниками многих биологически активных веществ, в том числе и гормонов [Крепс, 1979; Tocher, 2003].
Проведен анализ содержания общего белка и полного липидного состава кормов №№ 1, 2, 3 различных фракций (A, B и C) для аквакультуры радужной форели; полученные результаты представлены в таблице 1. Фракции корма № 1 отличались от соответствующих фракций других исследованных кормов высоким уровнем белка, холестерина, фосфати-дилхолина, насыщенных жирных кислот, о9 полиненасыщенных жирных кислот (ПНЖК) (см. табл. 1). Для комбикорма № 2, по сравнению с другими исследованными комбикормами, характерна более высокая концентрация триа-цилглицеринов (ТАГ), лизофосфатидилхолина, сфингомиелина, об ПНЖК и низкая - холестерина (ХС) и о3 ПНЖК (см. табл. 1). Отличительной особенностью комбикорма № 3 являлось высокое содержание фосфатидилэтаноламина (см. табл. 1).
В данном исследовании установлены различия не только между кормами разных производителей, но и между фракциями одного комбикорма, однако последние были менее выражены (см. табл. 1). Во всех исследованных комбикормах уровень ТАГ увеличился в ряду фракций кормов А-В-С, а концентрация холестерина (ХС), напротив, снижалась (см. табл. 1). При производстве различных фракций кормов учитываются возрастные особенности метаболизма лососевых. У молоди форели активно протекает пластический обмен, поэтому при изготовлении кормов для данной группы рыб используют источники сырья животного происхождения с высоким уровнем структурных компонентов (белок, холестерин, фосфолипиды (ФЛ)). В то время как для возрастных особей характерно накопление липидов в тканях и для
Таблица 1. Содержание белка (% сухой массы), липидных компонентов (% сухой массы) и некоторых жирных кислот общих липидов (% от суммы жирных кислот) в комбикормах
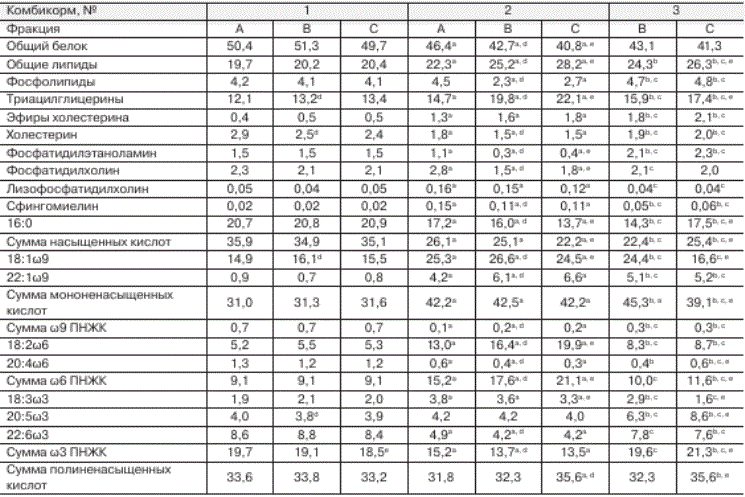
Примечание. а Различия достоверны при р < 0,05 при сравнении кормов 1 и 2 соответствующих фракций; ь различия достоверны при р < 0,05 при сравнении кормов 1 и 3 соответствующих фракций; с различия достоверны при р < 0,05 при сравнении кормов 2 и 3 соответствующих фракций; 11 различия достоверны при р < 0,05 при сравнении фракций А и В одного корма; е различия достоверны при р < 0,05 при сравнении фракций В и С одного корма.
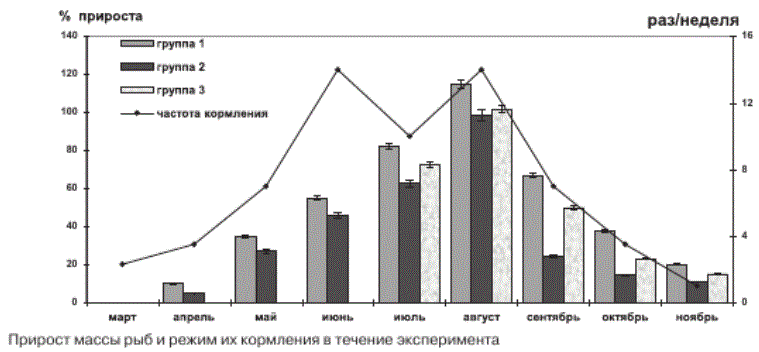
Прирост массы рыб и режим их кормления в течение эксперимента
них выпускают более калорийные комбикорма, включающие в свой состав растительные масла [Остроумова, 2001; Щербина, Гамыгин, 2006].
Низкий уровень структурных липидов (ФЛ и ХС) в корме № 2 (фракции В и С) по сравнению с аналогичными фракциями других изученных комбикормов (см. табл. 1) может быть связан с их длительным периодом хранения (более четырех месяцев). Ранее в наших исследованиях было показано [Назарова и др., 2010, 2013;
Таблица 2. Параметры роста радужной форели и смертность рыб
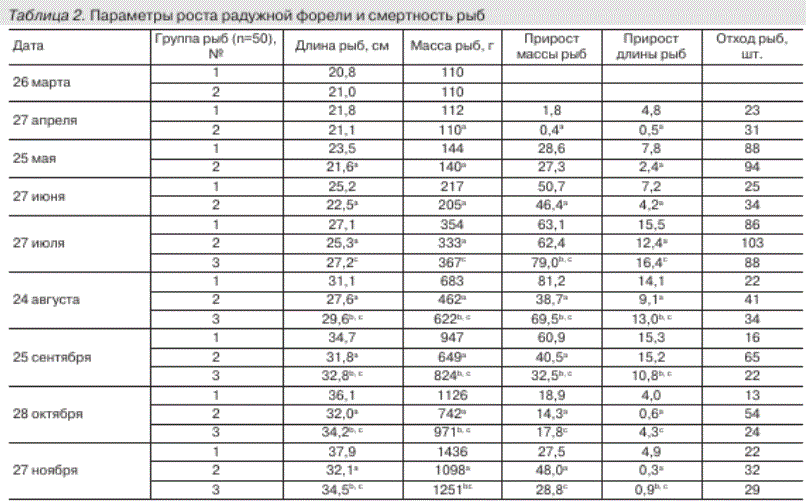
Примечание. а Различия достоверны при р < 0,05 при сравнении рыб групп № 1 и № 2; ь различия достоверны при р < 0,05 при сравнении групп рыб № 1 и № 3; с различия достоверны при р < 0,05 при сравнении групп рыб № 2 и № 3.
Немова и др., 2011], что, несмотря на срок годности продукта, указанный производителями комбикормов, составляющий шесть и более месяцев, уровень общих липидов и их отдельных фракций в кормах уже после четырех месяцев хранения заметно снижался. Содержание триацилглицеринов при хранении значительно не изменялось, сохраняя общую калорийность кормов. Наибольшей деструкции подвергались фосфолипиды комбикормов, что приводит к дефициту данного класса соединений в тканях форели и, как следствие, к снижению темпов роста рыб и ухудшению качества рыбной продукции для потребителя.
Ростовые процессы у гидробионтов зависят от целого комплекса внешних и внутренних факторов, среди которых одним из наиболее значимых является трофический, поскольку состав пищи и степень ее доступности во многом определяет линейно-весовую разнокачест-венность рыб [Дгебуадзе, 2001]. Трофические цепи гидробионтов формировались в течение длительного периода времени в процессе коэволюции, поэтому кормовая база рыб из природных водоемов имеет оптимальный химический состав для консументов [Pratoomyot et а1., 2011]. В условиях аквакультуры форель выращивают на искусственных комбикормах, исходное сырье для производства которых
должно максимально соответствовать естественной пище рыб. Введение в состав корма нехарактерных для натурального питания радужной форели компонентов может оказать значительное воздействие на метаболизм рыб и, как следствие, привести к изменению их физиологического состояния и ростовых процессов [Asian et al., 1996; Blanchet et al., 2005; Zaman et al., 2008].
Период исследования данной работы охватывал несколько этапов годового цикла рыб -переход от зимовального периода (март-май) к нагульному (июнь-сентябрь) и вновь к зимовке (октябрь-ноябрь). Пищевое поведение рыб определялось сменой режима кормления, включающего в себя частоту подачи корма и его количество, которое было одинаково для всех трех групп форели (рис.). Абсолютное количество корма, вносимого в садки, постепенно увеличивалось с марта по июнь, а в июле, в связи с высокой температурой воды, кормление рыб было снижено (см. рис.).
Динамика прироста массы форели между изученными группами была одинакова и определялась режимом кормления - при переходе с зимовального периода на нагульный темпы прироста массы рыб возрастали, а с сентября - снижались. Однако между группами рыб прирост массы форели различался (см. рис.) и зависел прежде всего от состава кормов, так как остальные условия культивирования рыб были одинаковыми. Темп роста рыб напрямую зависел от количества белка в корме. Наибольшее содержание общего белка было установлено в корме № 1 (см. табл. 1). Прирост длины и массы рыб, которые культивировались на данном комбикорме, был выше по сравнению с группами рыб № 2 и № 3 (табл. 2; рис.). Однако для активного роста рыб не менее важна и липидная составляющая корма, содержание которой должно быть сбалансировано, поскольку сравнительно высокий уровень липидов приводит к ожирению рыб, а дефицит отрицательно сказывается на их росте и развитии [Carter, 2003; Gümüs, ikiz, 2009]. Оптимальным для лососевых рыб возраста 1+ считается содержание липидов в корме около 20 % сухой массы [Остроумова, 2001]. В кормах № 2 и № 3 уровень общих липидов несколько превышал данное значение (см. табл. 1). ^оме количества липидов в корме важную роль играет и их качественный состав. Наибольшее значение имеет уровень структурных липидов (ФЛ и ХС) в корме, поскольку от их содержания зависит интенсивность пластического обмена. Фосфолипиды и холестерин формируют биомембраны, выполняя не только структурообразующую функцию, но и участвуя в регуляции активности мембраносвязан-ных ферментов [Hochachka, Somero, 2002]; ХС также служит предшественником стероидных гормонов и других веществ, необходимых для развития организма [Tocher et al., 2008]. Высокий уровень структурных компонентов в кормах № 1 и № 3 (см. табл. 1) способствовал более активному приросту массы рыб соответствующих групп.
На темп роста радужной форели влияет не только уровень ФЛ в корме, но и входящие в состав липидов полиненасыщенные жирные кислоты. ПHЖK играют важную роль в адаптации мембранных структур клеток эктотермных организмов к изменениям условий внешней среды, являются предшественниками биологически активных соединений, таких как эй-козаноиды, и выполняют ряд других важных функций [Hochachka, Somero, 2002; Bell et al., 2006]. В организме радужной форели осуществляется элонгация и десатурация жирных кислот, поэтому ПHЖK могут синтезироваться из незаменимых кислот - линолевой 18:2о6 и ли-ноленовой 18:3о3, содержание которых в исследованных комбикормах соответствовало необходимому, согласно некоторым рекомендациям, уровню [Остроумова, 2001; Blanchard, 2008]. Однако собственный синтез ПHЖK в организме рыб не восполняет их физиологически необходимое количество [Thanuthong et al., 2011], поэтому для нормального развития радужной форели необходим высокий уровень длинноцепочечных ПНЖК семейства о3 (таких как эйкозапентаеновая 20:5о3 и докозагексае-новая 22:6о3 кислоты) в комбикорме, которые способствуют активному росту рыб [Bell et al., 2010]. Соответственно, меньший уровень длинноцепочечных ПНЖК в корме № 2, по сравнению с другими кормами, возможно, определял более низкий темп прироста длины и массы форели соответствующей группы (см. табл. 2). Недостаток поступления ПНЖК в организм рыб может послужить причиной не только снижения ростовых процессов форели, но и нарушения функциональной активности органов и тканей рыб, вплоть до гибели организма [Carter, 2003; Grisdale-Helland et al., 2008].
Одним из индикаторов состояния популяции рыб является показатель их смертности. Больший отход рыб групп № 2 и № 3 по сравнению с группой № 1, возможно, связан с высокой долей эруковой 22:1о9 кислоты в кормах № 2 и № 3 (см. табл. 1 и 2). В настоящее время вопрос о влиянии высокой концентрации эруковой кислоты в комбикорме на метаболизм радужной форели является дискуссионным. По мнению одной группы авторов [Collins et al., 2012; Randall et al., 2013], эруковая кислота не оказывает значимого воздействия на физиолого-биохимические характеристики состояния рыб. Согласно другим литературным источникам [Sahasrabudhe, 1977; Tucker, Hargreaves, 2004], длительное употребление кормов, содержащих более 3 % эруковой кислоты от суммы жирных кислот, может привести к нарушению процессов роста и развития рыб и их гибели. Также на уровень смертности форели, возможно, могло повлиять и повышенное содержание ТАГ в комбикормах № 2 и № 3 (см. табл. 1 и 2), поскольку потребление кормов с высоким уровнем триацилглицеринов (более 20 % сухой массы) приводит к угнетению иммунной системы рыб и их физиологического состояния [Kj^r et а!., 2009].
Заключение
Комбикорма для лососевых различаются между собой соотношением структурных и запасных веществ, которое зависит от исходного сырья, используемого при производстве корма. В данной работе показано, что доля фос-фолипидов, холестерина и полиненасыщенных жирных кислот в комбикормах одного производителя снижается с увеличением крупки корма.
Динамика прироста длины и массы радужной форели зависит от режима кормления рыб. Показаны различия в приросте массы и темпах роста у трех групп рыб за изученный период, которые обусловлены, вероятно, составом комбикормов, на которых они культивировались. Качество кормов определяет темп прироста длины и массы рыб, интенсивность которых зависит от содержания белка, структурных липидов и о3 полиненасыщенных жирных кислот в комбикормах.
О. Б. Васильева1, М. А. Назарова2, П. О. Рипатти1 , Н. Н. Немова1
1 Институт биологии Карельского научного центра РАН
2 Вологодский государственный университет