
Распределение, питание и рост искусственно выращенной молоди семги (Salmo salar L.) после выпуска на участки рек с разными абиотическими условиями
Как показали результаты многолетних наблюдений, низкая выживаемость молоди семги, выращенной в условиях рыбоводных предприятий Мурманской области, после выпуска ее в естественные условия зависит от ряда причин: существенные физиологические отклонения, связанные с неудовлетворительными условиями содержания [1; 2 и др.]; патологии преимущественно алиментарного характера, приобретенные в процессе выращивания [3]; выработанное в условиях завода неадекватное речным условиям поведение [4; 5]; воздействие хищников; неправильный выбор мест и сроков выпуска [6; 7 и др.].
В настоящее время благодаря выпуску молоди в раннем возрасте (годовики вместо трехлеток) и переходу на качественные корма удалось добиться значительного улучшения физиологического состояния молоди [5]. Выпуск весной под лед существенно уменьшает массовую гибель молоди от хищников, неактивных в данное время. Низкий уровень воды до весеннего паводка дает молоди возможность закрепиться на индивидуальных участках.
Проблема этим не исчерпывается, и для понимания причин повышенной смертности заводской молоди после ее выпуска большое значение имеет исследование существенных факторов среды, с воздействием которых сталкивается привыкшая к заводским условиям содержания молодь лосося.
При выборе выростных участков для расселения рыбоводной продукции учитывается состояние естественного воспроизводства. К зарыблению рекомендуются те участки, на которых в результате маршрутной съемки в летний сезон обнаружены низкие или нулевые плотности расселения молоди семги естественного происхождения. С учетом недостаточно развитой инфраструктуры на практике выбор мест и объемов выпуска заводских годовиков семги зачастую диктуется наличием подъездных путей к этим участкам, необходимых для автотранспорта или снегоходов.
Для формирования объективных научных критериев, предъявляемых к местам выпуска молоди атлантического лосося, были осуществлены исследования с целью изучения воздействия абиотических факторов на адаптируемых годовиков. К этим факторам относится совокупность гидрологических характеристик выростных участков.
В процессе анализа успешности адаптации молоди к жизни на участках рек с разнообразными характеристиками были использованы показатели питания (количественные и качественные), темпов роста и распределения на выростных участках.
Материалы и методы
Материалом для работы служила молодь атлантического лосося (семги), выращенная в условиях рыбоводных заводов до возраста годовика (1.) и выпущенная в реку Умба бассейна Белого моря и реки Кола, Средняя и Акким (приток реки Тулома) бассейна Баренцева моря на нерестово-выростные участки (НВУ) в середине апреля - начале июня в 2013-2015 гг. В реках Кола и Умба существуют естественные популяции атлантического лосося, состояние воспроизводства которых оценивается как напряженное; в этой связи осуществляются ежегодные выпуски заводской молоди. В реках Средняя и Акким зарыбляемые НВУ недоступны для естественных популяций семги по разным причинам. Выпуск искусственно выращенных годовиков на эти участки производится в рамках компенсационных мероприятий.
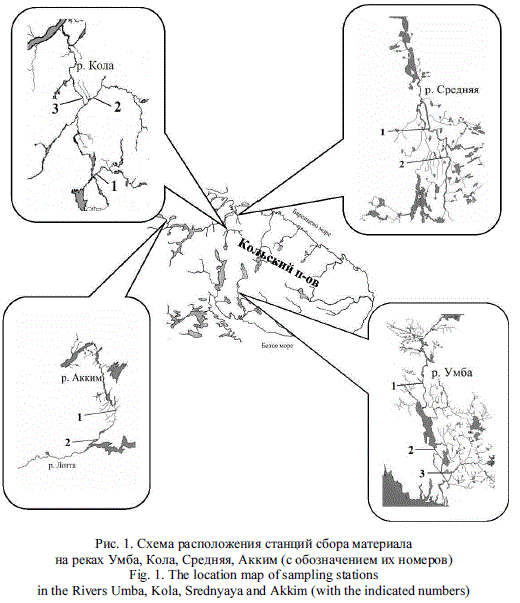
Контрольный облов рыб и сбор материала выполнялся через 1-5 месяцев в августе - сентябре в ходе маршрутной съемки с помощью специализированного электроловильного аппарата в разных биотопах в основном течении рек и в притоках (рис. 1). Молодь заводского происхождения идентифицировалась по отсутствию жирового плавника, ампутация которого является способом мечения молоди и осуществляется на всех рыбоводных предприятиях Мурманской области за 4-5 месяцев до ее выпуска. Оценивалось качество НВУ, различающихся по своим гидрологическим характеристикам (скорости течения, глубине, фракционному составу грунта), с помощью шкалы, предложенной для оценки грунта на НВУ [8]: П - песок (до 2,5 мм); ГМ - галька мелкая (2,5-5,0 мм); ГС - галька средняя (5-10 мм); ГК - галька крупная (10-25 мм); ВМ, ВС, ВК - валун мелкий (25-50 мм), средний (50-100 мм) и крупный (10-50 см); Г - глыбы. Грунт различных фракций группировался с присвоением баллов [9]. Наличие и степень обрастаний оценивали визуально и выражали в баллах: 0 - отсутствие обрастаний; 1 - единичные обрастания; 2 и 3 - слабая и средняя степень; 4 и 5 - сильное и сплошное обрастание, покрывающее 100 % дна. Использовались усредненные месячные данные по температурам воды в реках за период между выпуском молоди и временем ее поимки.
Расчет плотности расселения молоди осуществляли методом удаления [10]; пойманную рыбу немедленно фиксировали 70%-м этанолом.
Одновременно производили отбор проб дрифта стандартной ловушкой (площадь рамки 0,5*0,2 м; мешок из капроновой сетки № 19); время экспозиции составляло 30 минут. Пробы дрифта также фиксировали 70%-м этанолом. При обработке материала использовали количественно-весовую методику [11]. Содержимое желудков взвешивали на торсионных весах для определения индекса наполнения желудка (ИНЖ). Организмы из желудков разбирали по группам и просчитывали. Определение беспозвоночных проводили с использованием стандартных методик [12]. Индексы пищевого сходства (СП) определяли по методике А. А. Шорыгина [13]. Обработку ихтиологических материалов проводили по стандартной методике [14]. Удельную скорость роста С вычисляли по формуле
C = ln - l0 / lav ,
где ln - размер рыбы в конечный момент времени; l0 - размер рыбы в начальный момент времени; lav - средняя длина рыб за рассматриваемый промежуток времени [15]. В качестве временного интервала принимался 1 месяц.
Результаты и обсуждение
Описание выростных участков. Выбор ВУ для исследования был продиктован существенными различиями их гидрологических характеристик (табл. 1).
В районе р. Кола исследовались участки, расположенные в притоках Орловка, Кица и в основном русле реки (станция Лопарская). Данные ВУ имеют различные глубины и скорости течения, но также и сходную черту - здесь практически отсутствует водная растительность. ВУ на р. Орловка имеет глубину 0,2-0,7 м и каменистое дно, загроможденное крупными валунами в несколько слоев; скорость течения в межень до 0,8 м/с, в паводок несколько выше. Дно р. Кица преимущественно гравийно-галечниковое с отдельными крупными валунами, в летнюю межень значительно выступающими из воды; скорость течения на стрежневых участках достигает 1,0 м/с, а в прибрежной зоне не превышает 0,4 м/с. Выростной участок в основном русле р. Кола близ станции Лопарская является наиболее широким и протяженным; в период летней межени на значительной прибрежной акватории отмечаются глубина 0,3-0,4 м и скорость течения 0,3-0,5 м/с. Это создает оптимальные условия для распределения молоди лосося.
Самый протяженный участок выпуска в р. Умба расположен в среднем течении реки в Семиверстном пороге, где средняя скорость течения около 0,6 м/с и глубина 0,3-1,5 м. Грунты представлены крупным и средним валуном с примесью гальки; обрастания на стрежне отсутствуют, в прибрежной зоне они довольно развиты и представлены зелеными и диатомовыми водорослями. В протоке Родвиньга участки для выпуска заводской молоди расположены в 500 м ниже истока этой реки из Канозера. Максимальная скорость течения здесь составляет не более 1,0 м/с, а глубина - до 3 м. Грунт состоит преимущественно из крупной гальки, мелкого и среднего валуна. Обрастания слабые на стрежневой части, но развиты в прибрежной мелководной зоне. В притоке Вяла рыбу выпускают в протяженные порожистые участки, расположенные в нижней трети реки. Дно состоит из мелкого и среднего валуна с примесью гальки; средняя глубина 0,4 м; скорость течения до 1,0 м/с. Обрастания из мха рода Fontinalis покрывают до 70 % поверхности дна.
В р. Средняя первый ВУ расположен на 21 км от устья в верхней части порога. Субстрат здесь представлен валунно-галечными фракциями с преобладанием мелкого валуна. Обрастания из фонтиналиса покрывают практически 100 % поверхности дна. Скорость течения значительно варьирует (от 0,3 м/с в прибрежной части до 1,1 м/с на стрежне). Выростной участок, находящийся у моста пересекающей реку автодороги, представляет собой короткий перекат длиной около 100 м. Субстрат состоит преимущественно из песка и мелкого галечника. Обрастания в виде нитчатых водорослей покрывают около 10 % площади дна. Скорость течения варьирует от 0,1 м/с в прибрежной части до 0,5 м/с на стрежне.
В р. Акким молодь выпускали в порожистый участок в районе автодорожного моста. Этот участок характеризовался большой глубиной (до 2,5 м), умеренным течением и высокой степенью обрастания грунта, состоящего из мелкого и среднего валуна. Молодь распространилась на 5 км вниз по течению вплоть до устья реки, где преобладают небольшие глубины и скорость течения, грунт состоит из крупных фракций, а различные обрастания покрывают 100 % дна.
Среднемесячная температура воды в реках составляла от 11,6 °С в р. Кола и ее притоках до 13,5 °С в р. Средняя, что укладывается в рамки обычного для изучаемых рек диапазона значений.
Гидрологические условия во многом определяют видовой состав населяющей тот или иной водоток фауны беспозвоночных - основу питания молоди семги. Согласно классификации литофильных сообществ макробентоса, предложенной М. В. Чертопрудом [16], биоценоз исследованных рек можно отнести к классам 3 и 4. Для таких водотоков характерны олигосапробные условия, слабое заиление дна, развитые обрастания из видов рода Fontinalis на камнях в сочетании с крупными размерами самих камней. В сообществе беспозвоночных преобладают фильтрующие ручейники и поденки, хирономид мало.
Распространение молоди по участку. Как правило, годовики семги выпускаются в мелководные полыньи у берега. Мальки сразу опускались на дно, распределялись в пространстве между крупными валунами в несколько слоев и ориентировались в разных направлениях. Спустя несколько минут часть рыб начинала проявлять активность, постепенно перемещаясь по направлению к стрежню. Спустя 10-15 минут после выпуска основная часть рыб отплывала от берега, ориентировалась против течения и начинала перемещаться вниз по реке, используя энергию потока. Через 20-25 минут вся молодь рассредоточивалась настолько, что ее нельзя было увидеть.
При выпуске в промоины на быстром течении пестрятки сразу подхватывались мощным потоком. Первые несколько секунд они были полностью дезориентированы, но очень скоро проявляли реореакцию, разворачиваясь головой против течения, и, активизируя локомоторную функцию, устремлялись ко дну, где пытались укрыться в пространствах между камнями. Примерно половина пестряток уходила в стороны от открытого участка под лед, в тень.
Уже через месяц при облове ряда участков рек Кола и Умба попадания молоди регистрировали в основном ниже по течению на всем протяжении ВУ, в том числе на противоположной от места выпуска стороне рек. Молодь распространялась и вверх по течению, но в меньшей степени. Плотность ее расселения была самой высокой непосредственно в месте выпуска (от 35 экз./100 м2 в Орловке и Кице до 60 экз./100 м2 в Вяле) и заметно снижалась по мере удаления от него. Большая часть пестряток была сосредоточена у берега, в местах с умеренным течением (0,15-0,3 м/с), укрываясь среди выступающих из воды прибрежных камней. В это время годовики распределялись мозаично, образуя группы из нескольких особей, что свидетельствует об отсутствии у них территориального поведения, характерного для дикой молоди семги [17].
Осенью, спустя 3-5 месяцев после выпуска, в исследуемых реках пестрятки были расселены агрегированно на всех обследованных ВУ, образуя плотность от 10 до 30 экз./100 м2. Они по-прежнему занимали в основном прибрежную акваторию с невысокой скоростью течения и малыми глубинами, но встречались и на быстром течении ближе к стрежню (в относительно небольшом количестве). Речная молодь лосося, напротив, чаще присутствовала на быстром течении, и единично - у берега.
Динамика расселения молоди была одинаковой для всех без исключения наблюдаемых нами выпусков. Этот процесс в общих чертах представляет собой скорее не пассивный дрейф, а планомерное перемещение с использованием энергии потока и короткими остановками для поиска укрытия в грунте. Гидрологические характеристики выбираемых пестрятками участков соответствуют тем условиям, к которым молодь привыкла на рыбоводном заводе: небольшая глубина и относительно низкая скорость течения.
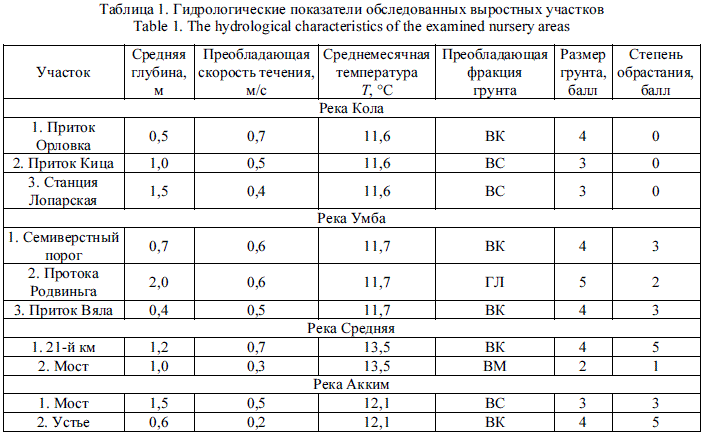
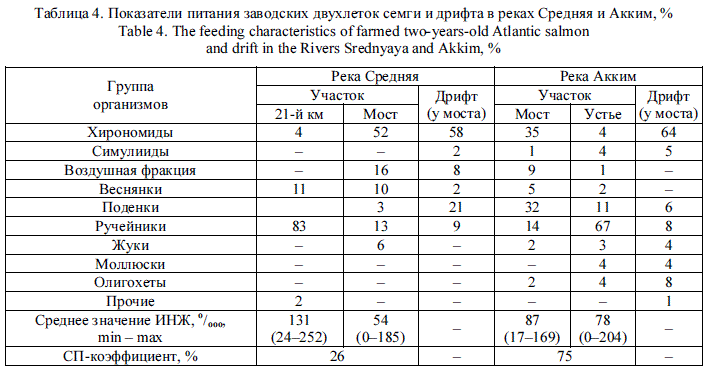
Питание молоди. Качественный состав питания заводских и речных пестряток на всех ВУ в р. Кола имеет существенные различия, но и та и другая молодь предпочитает личинок веснянок, поденок и ручейников (табл. 2). Несмотря на большую долю хирономид в дрифте, в питании пестряток эти организмы отсутствовали. Сходство питания естественной и заводской молоди более или менее выражено только на ВУ в районе станции Лопарская (СП-коэффициент равен 59 %).
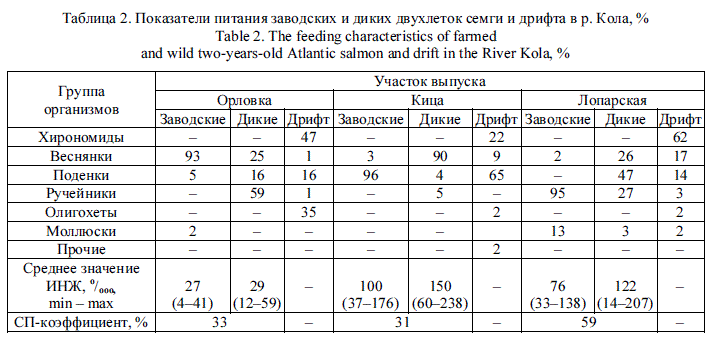
В р. Умба спектры питания заводской и речной молоди сильно различались на разных ВУ. Если в питании дикой молоди присутствовали 4—5 таксонов беспозвоночных, то пестрятки заводского происхождения употребляли в пищу представителей 5-9 таксономических групп. В основном пестрятки естественного происхождения отдают предпочтение личинкам ручейников и поденок, реже - личинкам симулиид. Определенной избирательности в кормлении у адаптируемых пестряток выявлено не было. Так, основу пищевого комка у заводских рыб на Семиверстном пороге составили моллюски, в протоке Родвинга -личинки ручейников, в Вяле - личинки веснянок, поденок и хирономид (табл. 3). СП-коэффициенты, оцениваемые на уровне отряда, во всех случаях оказались меньше 50 %.
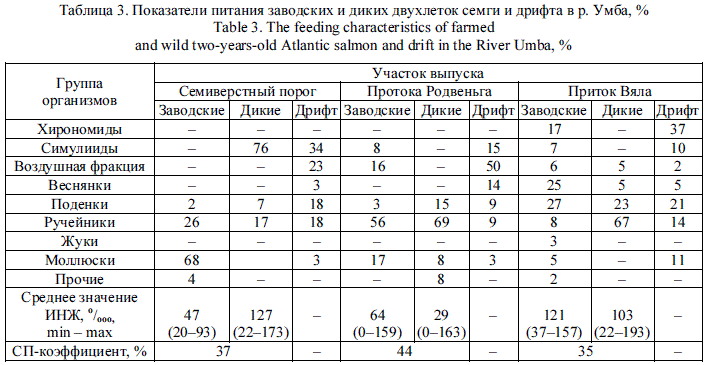
Наиболее разнообразный спектр питания наблюдался у молоди в реках Средняя и Акким (табл. 4). Например, в р. Акким в районе устья в пищевых комках молоди были обнаружены представители девяти таксономических групп беспозвоночных, а на участке около моста - восьми групп. СП-коэффициент был самым высоким - 75 %.
Индексы наполнения желудков у рыб на всех ВУ характеризовались широким хаотическим разбросом значений, достоверно не различаясь у естественной и заводской молоди (табл. 2-4). Анализ проб питания в ряде случаев может показать более высокий индекс наполнения желудка у естественной молоди, чем у заводской [18], либо наоборот [17]. В нашем случае сопоставление этого показателя с факторами среды не выявило тесных математических зависимостей, что было ожидаемо, поскольку интенсивность питания -величина непостоянная и зависит в большей степени от сезонной и суточной динамики дрифта, его плотности, различий в микросреде обитания и других факторов [19-21]. Воздействие низких температур проявлялось в первые недели адаптации: показатель ИНЖ заводских годовиков в это время на порядок ниже, чем у их диких сверстников. Впоследствии интенсивность питания на фоне общей активности годовиков увеличивалась по мере роста температуры окружающей среды [17].
Следует отметить, что отлов молоди осуществлялся нами при температурах воды от минимальных значений (0,1 °С в апреле и 2,5 °С в сентябре) до максимальной температуры (15,5 °С в августе) со значительно различающимися гидрологическими показателями. Во всех случаях желудки молоди содержали те или иные пищевые организмы, что подтверждает мнение ряда специалистов об "универсальности" ювенильного лосося, способного успешно питаться в разных местах обитания (от небольших ручьев до крупных озер) и в различных условиях [20]. При необходимости молодь может совершать миграции на участки с лучшей кормовой базой, например, в небольшие притоки из основного русла [22]. Рост молоди. Коэффициент удельной скорости роста заводской молоди продемонстрировал значительную вариабельность. Если в р. Умба он был относительно стабилен, то в остальных реках темп роста пестряток на разных ВУ отличался в разы (табл. 5). Это наводит на мысль о наличии неких факторов, способных существенно модифицировать темп развития пестрянок.
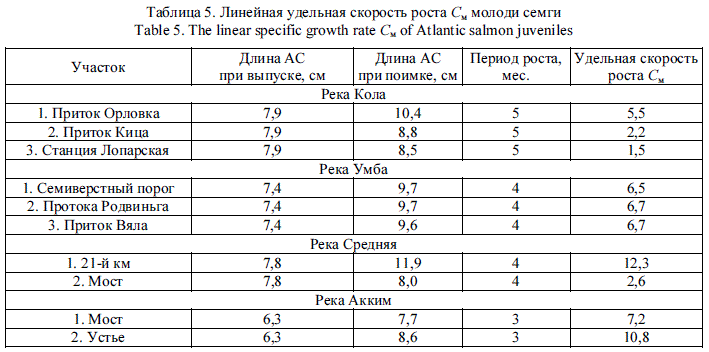
В отличие от вариабельного показателя индекса наполнения желудка, показатель роста молоди является наилучшим индикатором успешности ее адаптации к естественным условиям. В норме поведение молоди направлено на максимальное снижение потери энергии и максимальное потребление пищи [8]. Те заводские пестрятки, которые выработают нормальное пищевое поведение, к осени наберут большие длину и массу и получат больше шансов на выживание в зимний период. Следовательно, темп роста можно рассматривать в качестве интегрального показателя, зависящего как от обилия кормового ресурса, так и от способности пестрятки добыть его с минимальными энергетическими затратами.
Для выявления абиотических факторов, способствующих развитию молоди, было проведено простое попарное сопоставление удельной скорости роста с показателями скорости течения, глубины, температуры, типом грунта и степенью донных обрастаний. Последние два показателя выражены в баллах (табл. 1, 5). Количество сопоставляемых пар значений - 10. Скорость роста адаптируемых пестряток статистически не зависит от скорости течения (Я = 0,16), глубины (Я = -0,01) и температуры (Я = 0,10), которые в исследуемых случаях укладывались в диапазон "норма" и не оказывали выраженного влияния. Обнаружены тесные положительные связи скорости роста с преобладающим размером грунта (Я = 0,70) и степенью донных обрастаний (Я = 0,88). Найденные связи выражены графически линейными регрессиями (рис. 2).
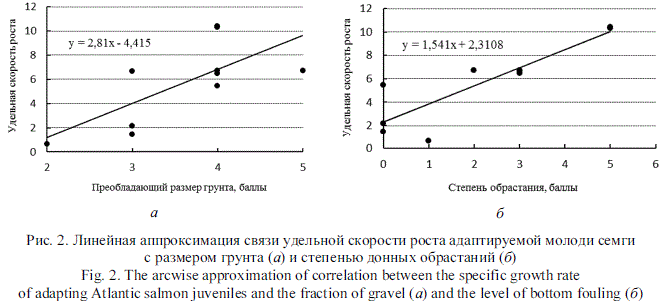
Полученный результат, на наш взгляд, следует рассматривать как отражение приспособительной реакции заводских пестряток к незнакомым условиям среды обитания. Начиная с момента выпуска пестрятки пытаются найти привычную для себя гидрологическую нишу, что приводит к их расселению в прибрежной зоне участка рек со слабым течением. Одновременно, следуя инстинкту, молодь занимает индивидуальные участки, о чем свидетельствуют переход от стайного к агрегированному характеру распределения и расселение по всей протяженности ВУ. Отсутствие у заводской молоди навыка добывать пищу из потока приводит к существенному отличию от речных сверстников в качественном составе питания. Следовательно, наилучшее сочетание факторов обитания для адаптируемой молоди, обеспечивающее быстрое накопление вещества и энергии, - малая скорость течения, наличие укрытий и достаточное количество легкодоступного корма. Лучше всего этим требованиям соответствуют прибрежные участки порогов со слабым течением и валунным грунтом, покрытым мхом, который служит как дополнительным укрытием, так и благоприятным субстратом для беспозвоночных. Одно из функциональных свойств грунта - формирование условий распределения рыб посредством образования топогидравлических ниш, размер и число которых зависят от сочетания различных фракций [8]. Кроме того, каменистое дно является субстратом для литореофильных биоценозов, включающих амфибиотические организмы - излюбленный корм молоди лосося.
Неприспособленность к жизни в естественной среде является результатом морфологических, физиологических, экологических и поведенческих изменений, произошедших в период жизни в условиях рыбоводного завода [1; 23]. Проблема адаптации заводской молоди к речным условиям изучается давно. В частности, доказано, что содержание рыбы в информационно обогащенной среде и применение живого корма перед выпуском может значительно улучшить показатели выживаемости после выпуска выращенных рыб [4; 24 и др.]. К сожалению, успешные результаты лабораторных экспериментов очень сложно внедрить в биотехнику выращивания. Корректируя условия выпуска посадочного материала, в том числе выбирая более подходящие участки для адаптации молоди, можно добиться увеличения выживаемости адаптируемой молоди и промыслового возврата при искусственном воспроизводстве.
Заключение
Исследование воздействия абиотических факторов на распределение, питание и рост в реках годовиков атлантического лосося, выращенных в условиях рыбоводных предприятий Мурманской области, проведено нами впервые. Начиная с 2006 г. рыбоводные заводы Мурманской области перешли на выпуск молоди в возрасте годовика, пытаясь минимизировать заметные недостатки развития, наблюдаемые при выращивании молоди до трехлетнего возраста.
Установлено, что расселение заводской молоди по всему протяжению выростных участков завершается к окончанию вегетационного сезона. Пестрятки рассредоточиваются в основном в местах со слабым течением у берега, занимая индивидуальные участки. Качественный состав их питания значительно отличается от рациона дикой молоди; он неодинаков у пестряток, обитающих на разных участках. Количественный состав питания адаптируемой молоди не зависит от глубины, скорости течения и других абиотических факторов.
Темп линейного роста положительно связан с размером фракций грунта дна реки и степенью развития водных обрастаний: наивысший темп линейного роста молоди отмечен на участках, где имеются камни крупных размеров в сочетании с сильной степенью обрастания дна макрофитами.
В качестве практической рекомендации представляется возможным предложить оптимизировать процесс выпуска, подбирая участки для расселения рыбоводной продукции не только исходя из оценок плотности речной молоди, но также учитывая структуру дна.
А. М. Николаев, М. Ю. Алексеев, И. В. Самохвалов, А. Г. Легун, Н. В. Ильмаст, Е. Н. Распутина, Ю. А. Шустов