
Морфофункциональные особености лососей
Близкое родство и принадлежность изучавшихся рыб к одной экологической группе обусловили их большое сходство. В то же время мы видели достаточно четко выраженное своеобразие абиотических и биотических условий нереста различных лососей, режиманерестовых бугров, образа жизни молоди, меняющегося в онтогенезе. Поскольку процесс адаптивной радиации, освоения различной среды закономерно связан с морфофункциональной специализацией, становится очевидной важность познания специфики системы видовых адаптаций во всем многообразии ее проявлений.
Продуцирование спермы, время сохранения спермиями фертильности. В условиях наших опытов самцы выделяли сперму от полумесяца (нерка) до месячного срока (кета). У отдельных самцов кеты и кижуча полноценную сперму удавалось получить более 10 раз и общее ее количество достигало, соответственно, более 130 и 190 мл, тогда как у нерки — менее 50 мл (табл. 18). Продолжительное продуцирование спермы стоит в связи с выметыванием икры небольшими порциями и участием самцов в нерссте с несколькими самками. В отгороженных участках нерестилищ нерки икра успешно оплодотворялась даже тогда, когда на одного самца приходилось до 15 самок (Mathisеп, 1962; Матиссн, 1963). Но в природе наблюдается иная картина: у каждой икромечущей. самки держится по нескольку самцов. Их обилие повышаст вероятность оплодотворения икры (Солдатов, 1912; Shuпап, 1950).
Привлекают внимание экологические зависимости. Для нерки типично большое число самцов в гнездовых группировках, высокая концентрация производителей и слабая проточность нерестилищ.
Та блнца 18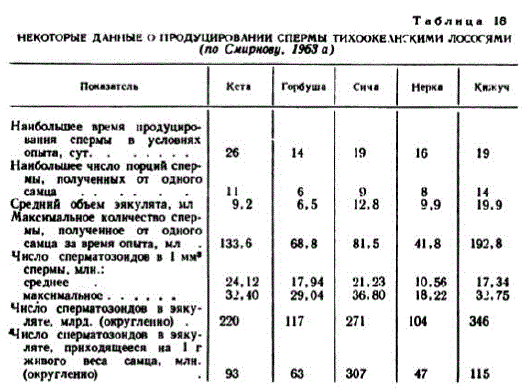
Сима, наоборот, нерестится на быстром течении, гнезда располагаются редко и около самки чаще держится один проходной и несколько карликовых самцов. Ее самцы продуцируют много густой спермы, а нерка выделяет мало спермы и она жидкая. В эякуляте нерки сперматозоидов в два с лишним раза меньше, нежели у симы и кеты, а на каждый грамм живого веса самца их приходится в 6 раз меньше, чем у симы (табл. 18). Интересен пример горбуши. Она нерестится на более быстром течении, нежели кета, но сперматозоидов продуцирует меньше. Как и у нерки, это компенсирустся большим числом самцов в гнездовых группировках и высокой плотностью заполнения нерестилищ. Таким образом, для успешного оплодотворения икры важны как обильное выделение спермы, так и высокая концентрация производи.6חTe
Сперматозоиды сохраняются в воде разное время: у лососевых и карповых рыб десятки секунд или несколько минут, у осетровых — несколько часов, у сельди и некоторых других морских рыб этот срок измеряется сутками (Гинзбург, 1968). У рассматpиваемых видов не обнаружено больших различий, но все-таки они имеются. В наших опытах через 20 сек. после разведения спермы водой у кеты в разных сериях оплодотворялось 75 и 90% икринок, у симы—до 90%, горбуши—60 и 70%, кижуча около 50%, а у нерки — лишь около 20% икринок. Через 60 сек. пребывания спермиев в воде у кеты оплодотворялось до 70% икринок, у симы — 60, у горбуши около 30%, у кижуча — единичные икринки, а спермии нерки полностью утрачивали фертильность. В отдельных опытах сперматозоиды горбуши и симы оплодотворяли единичные икринки после пребывания в воде в течение 2,5—3 мин. Сперма чавычи, разведенная водой, сохраняет фертильность до 5 мин. (Rutter, 1904). В отдельных опытах Ферстера (1968) некоторое количество икринок нерки оплодотворялось после того, как сперматозоиды пробыли в воде, температура которой менялась от 2 до 8°, 1 и даже 8 час. В других опытах разведенная сперма персставала оплодотворять икру уже через несколько минут. Какие причины могли дать столь колеблющиеся результаты не совсем ясно. Поскольку в опытах температура снижалась до 2°, можно предположить, что консервирующее действие оказывало OXHCHHC,
Особенности икры тихоокеанских лососей. Среди костистых рыб нашей фауны тихоокеанские лососи выметывают самые крупные икринки. Этот признак подчеркивал Гюнтер (1886) и использовал в качестве таксономического при обосновании самостоятельности рода Опсоrhynchus, представители которого ранее включались в род Salпо. Следует, однако, отметить существенные видовые вариации. Икринки чавычи, ксты, симы действительно выделяются крупным размером, у других видов они мельче, а нерка по этому признаку стоит в одном ряду с представителями рода Salпо.
Икринки лососей олигоплазматические. Количество цитоплазмы у разных видов колеблется незначительно. Вариации размера икринок определяются количеством дейтоплазмы. Это ведет к существенным различиям соотношения цитоплазмы и желтка. Например, в икринках диаметром около 6 мм, встречающихся у перки, кижуча и горбуши, желтка примерно в 400—500 раз больше, чем цитоплазмы, а в икринках диаметром 8 мм, свойственных кете и чавыче, это соотношение возрастает до 1000.
Очевидный выигрыш откладки икринок с обильным запасом питательных веществ, как уже говорилось, состоит в том, что потомство получает возможность длительный срок использовать защитные свойства нерестовых бугров, достигать в них крупного размера, высокого уровня дифференцировки и активности, благодаря чему в дальнейшем увеличиваются шансы ухода личинок от хищников. Кроме того, у крупных личинок рыб потребность в пище относительно меньшая, а энергия плавания больше. В результате расширяются районы поисков пищи (Marshall, 1963). Это особенно выгодно в условиях малой кормности нерестово-вырастных водомов,
Отмечена положительная связь между длиной самок и размерами икринок у осенней кеты Амура (Бирман, 1952). Указывается на общую тенденцию увеличения икринок с возрастанием размера лососей (Rounsefell, 1957). Однако картина довольно пестрая. Из тихоокеанских лососей наиболее мелкую икру выметывают нерка и кижуч, которые превосходят своими размерами горбушу и сахалинскую симу. У горбуши, самого мелкоголосося, икра довольно крупная. Средний по размеру лосось — кета, выметывает очень крупные икринки, сходные по размерам с икринками чавычи — наиболее крупноголосося. Уместно упомянуть и об относительно мелкой икре благородных лососей, достигающих большой длины H BeCo.
В литературс указывается на увеличение размера икринок с продвижением рыб к северу (Расс, 1947, 1953; Marshall, 1953). При сравнении видов и внутривидовых форм мы встречаемся с фактами иного рода. Сима, например, занимает южную часть ареала рода, но выметывает значительно более крупную икру, нежели нерка и кижуч, размножающиеся севернее. Крупным размером икринок выделястся осенняя кета южносахалинской р. Найбы. Чавыча камчатской р. Паратунки выметывает икринки весом около 240 мг, тогда как всс икринок чавычи из р. Кламатч, расположенной на юге ареала вида, в полтора раза больше (Leitritz, 1960). Привлекает внимание интереснейший факт уменьшения размера икры у горбуши, перессленной из южных рск Сахалина и Южных Курил в северные баренцевоморские и беломорские реки (Персов, 1963; Галкина, 1965; Сакун, 1965; Смирнов и др., 1968). Подобные факты стимулировали поиски экологической обусловленности размера икры лососей. Было логично сопоставить сезон, условия нереста и развития и размер икринок. При этом выявилась достаточно определенная связь величины икринок с термическими условиями нереста и развития (Смирнов и др., 1968). ፻፪ оказалось, икра крупнее и содержит больше питательных веществу лососей, нерестящихся и развивающихся при более высокой температуре. Этим обеспечиваются быстрый темп развития зародышей и их повышенная двигательная активность. Рассмотрим данные о составе икры. Икринки тихоокеанских лососей крупнее и богаче питательными веществами, нежсли у благородных лососей (табл. 1). Среди представителей рода Опсогhynchus крупным размером и повышенным содержанием сухого вещества (более 44%) выделяется икракеты и симы; учитывая данные Грина (Greene, 1926), то же следует сказать и об икре чавычи. Икра указанных видов и горбуши содержит больше жира. В сравнении с ними у кижуча жира примерно вдвое меньше. Несколько пониженным содержанием протеина выделяются икринки нерки. По физиологическим тепловым эквивалентам протеина и жира был проведен ориентировочный подсчет энергетичcской обеспеченности зародышей. Полученные показатели несколько занижены, поскольку не учитывались углеводы, которых в икре немного, однако они полностью сохраняют сравнительное значение (табл. 19). По этому показателю, как и некоторым другим, нерка сближается с атлантическими лососями. Все виды с большим запасом тепловой энергии в икринках, кроме осенней кеты, нерсстятся летом при сравнительно высоких температурах. Осенняя же формакеты, изучавшаяся нами, как и осенненерестующая чавыча. мечет икру в мсстах выхода теплых грунтовых вод и длительное время развивается при температуре 8—10°. Примечательно, что на нерестилищах Теплого озера в бассейне Амура вода холоднее (около 3—5°) и нерестящаяся здесь осенняя кета выметывает более мелкую икру.
Возникает вопрос, не противоречит ли нашим построениям тот факт, что нерка откладывает мелкую и бедную питательными веществами икру, а нерестится в августе, в теплое время года? Оказывается нет. Дело в том, что она устраивает гнезда на мсстах выхода грунтовых вод, сохраняющих летом низкую температуру. По размеру и запасам питательных веществ к икре нерки близка икра кижуча. Этот вид нерестится осенью и зимой. Семга из реки Кола и балтийский лосось нерестятся поздно и развиваются также при низкой температуре (Новиков, 1953). Так как некоторые виды и расы лососей нерестятся на местах выхода грунтовых вод, температура которых не связана с широтой местности, географический параллелизм в изменении размера икринок этих рыб отсутствует.
Икра лососей имеет яркую окраску, обусловленную высокой концентрацией каротиноидных пигментов. В икре и мышцах видов рода Oncorhynchus содержится единственный из каротиноидов ксантофиллов — астаксантин. У представителей рода Salпо иместся астаксантин, а также лютеин и зеаксантин (Cilover, Morton, Rosen, 1952; TyBHH, 1953; Kanemitsu, Aloc, 1958; Slip»KoMбек, 1964, 1970, 1972; Яржомбек, Грачев, 1964; Боровик, 1966; Ирискина, 1968; Приходская, 1972). Каротиноиды чpcзвычайно широко распространены в различных органах и тканях растений и животных, выявлено большое количество этих пигментов, им приписываются разнообразные функции (Дунаевский, 1947; Будницкая, 1952; Лебедев, 1953; Гудвин, 1954; Балаховский, Дроздова, 1956; Еремеев, 1957; Яржомбек, 1970; Карнаухов, 1971, 1973). Поскольку лососи развиваются под покровом грунта, окраска икры и зародышей не может иметь криптического или сигнального значения. Равным образом и при сосредоточении во внутренних органах каротиноиды не могут выполнять присущей им фоторсцепторной или фотосинтетической функций. Высказывалось предположение об участии каротиноидов в процессе оплодотворения, но оно не получило подтверждения (Гудвин, 1954). В то же время трудно согласиться с отрицанием функциональной нагрузки у этих биологически активных веществ в период эмбрионального развития лососей и других рыб или с признанием важности пигмента яиц в первую очередь для формирования хромотофоров у будущих мальков (Steven, 1948). Широкое распространение каротиноидов в природе и указания на их разнообразную роль в организме наводят на мысль, как справедливо заметил Т. Гудвин (1954), о допустимости некоего общего фактора или свойства, «посредством которого все эти функции являются связанными».
При поисках ответа на вопрос о значении разнообразия окраски икры и зародышей рыб нами было обращено внимание на связь интенсивности пигментации с кислородными условиями среды обитания. Сравнительный эколого-эмбриологический анализ показал, что икра, покровы и некоторые внутренние органы зародышей интенсивнее пигментированы у фитофильных, остракофильных, вынашивающих и некоторых живородящих видов, а также
рыб, прячущих икру под камни, ракушки и в грунт (Смирнов,
1947, 1950; Крыжановский, 1948, 1949). Представители указанных экологических групп развиваются в среде, обедненной кислородом. Наоборот, у видов, развивающихся в лучше аэрированной воде (пелагофильных, псаммофильных), икра и зародыши пигментированы слабо. Наблюдается следующий параллелизм: у видов. имеющих интенсивно окрашенную икру и зародышей, эмбриональные органы дыхания развиваются раньше и мощнее, а в случаях слабой пигментации они формируются позже, наблюдастся редукция сосудистой системы. Установленные эколого-морфологические зависимости и известные в то время суждения о возможном участии каротиноидов в окислительно-восстановительных процессах позволили высказать предположение о том, что каротиноидные пигменты функционально дополняют сосудистую дыхательную систему рыб. Представление об экологической обусловленности каротиноидной пигментации было распространено на объяснение разнообразия окраски взрослых рыб, в том числе лососей в брачную пору (Смирнов, 1950, 1954a, 1959б).
Связь интенсивности пигментации икры и зародышей, равно как и развития эмбрионально-личиночных органов дыхания, с условиями аэрации среды обитания в дальнейшем была подтверждена при эколого-эмбриологическом изучении рыб сем. Gasteгоsteidae, Cottidae, Cottocomeforidae, Clupeidae, Percoidei, Jococeň родов Salmo и Опсогhynchus (Крыжановский и др., 1953; Смирнов, 1953б, 1954a, 1955а, 1958, 1959б, 1960а, б, 1964а, б; Соин, 1954, 1956, 1964, 1968; Крыжановский, 1956).
Сопоставление пигментации икры тихоокеанских лососей, развития эмбриональных органов дыхания и условий аэрации гнcзд дает достаточностройную картину. Так, икра горбуши выделяется светлой оранжевой окраской, небольшим количеством астаксантина; эмбриональные органы дыхания у нее развиты слабее родственных видов и развивается она при высоком насыщении воды кислородом. Икринки кеты ярко-оранжевые, содержат больше каротиноидов, эмбриональные органы дыхания у нее развиты мощнее горбуши, эмбриогенез кеты протекает в условиях меньшего содержания кислорода. У других видов икра окрашена интенсивнес и на желточном мешке развивается особенно густая сосудистая сеть. Наиболее богата астаксантином икра нерки (Яржомбек, 1970), сосудистая сеть у ее эмбрионов развивается быстрон мощно, нерест этого вида приурочен к местам выхода грунтовых вод, бедных кислородом (Крохин, 1960).
271
Заслуживают внимания данные физиологических опытов, в которых отмечена повышенная стойкость к высокой температуре более интенсивно окрашенной икры свирского сига, по сравнению с окрашенной слабее. У той же икры в некоторых опытах отмечался повышенный окислительный обмен, что связывалось с обилием в икре витамина А (Баранчесв, Разумовский, 1937). Большая устойчивость интенсивно пигментированной икры к неблагоприятным внешним факторам наблюдалась у лососей и других рыб (Hubbs, Stevenhagen, 1958; Логинова, 1966, 1969; Шнаревич, Сахненко, 1968; Яржомбек, 1970; Приходская, 1972). Интересно, что при инкубации икры радужной форсли в воде с низким содержанием кислорода выявлена прямая зависимость между количеством каротиноидов в икре и интенсивностью дыхания эмбрионов, тогда как при нормальном насыщении воды кислородом подобной корелляции не обнаруживалось (Бровик, 1966). Сообщается о положительном влиянии каротиноидов на рост, развитие и выживаемость эмбрионов птиц
(Еремеев, 1957).
Проблема связи каротиноидов с окислительным метаболизмом разрабатывастся со времени публикаций К. С. Мерсжковского (1883) H M. ApHo (Arnoud, 1886; u HT. no E5yLHH LI KorA, 1952).: M3 BecTна гипотеза А. Н. Баха (1912) об участии этих соединений в качестве оксигеназ при ферментативном окислении, получившая подтверждение в модельных опытах (Бородина, 1939). Показана способность каротиноидов и близких им веществ активировать молекулярный и перекисный кислород; утверждастся, что понимание роли каротиноидов в деятельности организма не представляется возможным без изучения их участия в окислительных процессах (Балаховский, Кузнецова, 1955; Балаховский, Дроздова, 1956). Имеются сведения об участии данных веществ в окислительно-восстановительных реакциях и их важной роли в приспособлении растений к неблагоприятным условиям существования (Лебедев, 1953). Исследустся функция каротиноидов в механизме переноса кислорода в процессе фотосинтеза (Сааков, 1964; Сапожников и др., 1964). Представления о роли каротиноидов в окислительном метаболизме основываются на наличии большого количества ненасыщенных сопряженных двойных связей и хороших электронно-донорных и электронно-акцепторных свойствах этих веществ. В результате анализа материалов, полученных при изучении каротиноидов в нервной и мышечной тканях и литературных данных В. Н. Карнаухов (1971, 1973) пришел к выводу о вероятном их участии совместно с гемопротеинами в процессах внутриклеточного депонирования кислорода, позволяющего компенсировать малую скорость его поступления в условиях гипоксии.
Обсуждая значение каротиноидов в приспособлениях лососей, связанных с процессами размножения и развития, нельзя обойти их роли в формировании брачного наряда. При этом следует иметь в виду не только приобретение производителями яркой окраски, оптический эффект, но и вероятное участие этих веществ в окислительном мстаболизме. Указывается также, что каротиноиды и близкие им вещества могут тормозить кератинизацию покровов, способствуют восстановлению эпителия, в культурах тканей изменяют покровные ткани в сторону уподобления их слизистому эпителию, повышают сопротивляемость организма инфекциям и т. д. (Балаховский, Кузнецова, 1955; и др). В подобных многосторонних влияниях каротиноидов может быть усмотрена их важная роль в структурных преобразованиях кожи лососей в период созревания, которые благоприятствуют интенсификации кожного дыхания и позволяют лососям во время нереста находиться во взмученной, обеднепной кислородом среде и выполнять работу по расчистке грунта и устройству нерестовых бугров (Смирнов, 1959 б).
Функции каротиноидов в процессе развития и жизнедеятельности организмов, в том числе рыб, как говорилось, разнообразны. С удовлетворением можно отметить, что эта сложная проблема привлекла внимание не только эмбриологов, но также физиологов, биохимиков, биофизиков, что позволяет надеяться на ее успешную дальнcйшую разработку.
Среди других особенностей икры привлекает внимание различис строения и свойств оболочек, выполняющих роль посредника между зародышем и внешней средой и несущих защитную функцию. После вымcтывания икриноких поверхность на короткос время приобретает слабую клейкость. Внешний гомогенный слой защищает яйцевые оболочки от разрушения протеолитическими бактериями и, вероятно, — от воздействия фермента, выделяемого при вылуплении зародышей из соседних икринок (Bell et al, 1969). Икринки лососей обладают высокой прочностью. Развивающиеся икринки форели и атлаптическоголосося могут выдержать давление 3—3,5 кг, улетней кеты — 4—5 кг, у горбуши—8—9 кг. Особая прочность икринок горбуши связывается с сснерестом на быстром течении, увеличивающем опасное для потомства поверхностное перемещение грунта (Никольский, Соин, 1954; Соин, 1954). Как видно из табл. 19, у этого вида оболочки наиболее толстые, их толщина уменьшается в ряду: кета, чавыча, кижуч, сима, а наиболее тонкие оболочки свойственны неркс. Последняя нерестится на участках сослабым течением или где оно отсутствуст, и наличие у икринок нерки тонких оболочек подкрепляст мнение о связи толщины оболочск со скоростью течения на нерсстилищах. Однако подобная зависимость не носит общего характера. Так, на быстром течении нерестятся сима и кижуч, но яйцевые оболочки у них довольно тонкие. Дело в том, что зародыши лососей от повреждений при поверхностных подвижках грунта могут быть защищены также глубокой закопкой икры. Консчно, закопать икру глубокомогут лишь более мощные рыбы, такие, как чавыча, кижуч, и у этих видов мы видим более тонкие оболочки, нежели у горбуши и кеты. Естественно также предположить, что виды, развивающиеся в воде бедной кислородом или при неустойчивом газовом режимс, особенно толстых оболочек иметь не могут. Это предположение подтвер
ждает пример нерки, кижуча и симы. Наоборот, горбуша развивается в условиях более высокого насыщения воды кислородом и поэтому смогла пойти по пути приобретения особенно толстых яйцевых оболочек.
Выявлены структурные отличия оболочск. Внутренний слойлучистой зоны яйцевых оболочек у кеты и горбуши выражен отчcтливее, его структура болес рыхлая, а у нерки на гистологических препаратах он выделяется лишь интенсивностью окраски. Различается и внешний аморфный слой оболочек. Икринки чавычи, например, имеют белесоватую поверхность, дающую в отраженном свете своеобразный перламутровый отблеск. Яйцевысоболочки у горбуши непрозрачны, а у икринок нерки и симы они прозрачнее оболочек других лососей.
После попадания в воду икринки сохраняют способность оплодотворяться разное время. Сообщается, что этот срок у икринок кеты достигает 15—30 мин. (К. Үаппапоto, 1951). Так долго могут сохранять фертильность лишь немногие икринки и в своеобразных условиях. В наших опытах через 1 мин. после помещения в воду теряло фертильность 50% икринок кеты, 44% горбуши и 15% икринок симы и почти все икринки утрачивали фертильность у кеты через 6 мин., у горбуши—через 7—8 мин., у симы--после 10 мин., крайние сроки сохранности отдельных икринок достигали 12— 15 мин. Приведенные данные указывают на необходимость строгого соблюдения в лососеводстве методики «сухого способа» осеменения икры, гарантирующего быстрый контакт икринок со сперMaTO3OHua M. H.
Периодизация онтогенеза тихоокеанских лососей и некоторые особенности прохождения отдельных периодов и этапов развития. В эмбриональном периоде развития осенней кеты, которые у лососей продолжителен, Н. Н. Дислер (1954, 1957) выделил 12 этапов, в личиночном — один и в мальковом периоде жизни — два этапа. Разработанная им периодизация онтогенеза тихоокеанских лососей принята многими авторами (Крыхтин, 1955, 1962; Смирнов, 1958, 1963 б; Васильев, 1959 б; Фроленко, 1966; Лукина, 1966. 1973). В настоящей работе за основу взята та же схема, в которую внесены некоторые уточнения.
За начало жизни особи мы приняли образование зиготы. С вылуплением связаны разносторонние изменения. Зародыш освобождается от оболочек и пернвитсллиновой жидкости. Он вступаст в непосредственный контакт с потоком, частицами грунта и сго отношения с внешней средой принципиально меняются, вследствие чего изменяется функциональная активность органов дыхания, движения, экскреции, органов чувств, то есть меняется вся биология. Поэтому мы полагаем правильным акт вылупления считать границей смежных этапов. Далее при описании развития обращалось вниманис на разнообразие уровня морфологической диффсренцировки, при котором молодь мигрирует в море. Видовые различия продолжительности пресноводного цикла жизни лососей подробнее будут обсуждены ниже. В данном случае имеется в виду подчеркнуть широкую внутривидовую и внутрипопуляционную вариабельность этого признака. Наглядный пример прсдставляетнерка. Этот лосось может мигрировать в море личинками, сеголетками, после одной или нескольких зимовок в пресной воде и, наконец, созревать без выхода в море. Подобные примеры свидетельствуют о том, что момент катадромной миграции не сопряжен с достижением строго опредсленного уровня дифференцировки и это затрудняет выделение особого этапа ската. В связи с периодизацией оптогенеза были рассмотрены также особенности брачного периода жизни лососей. Каждого наблюдателя поражают глубокие преднерестовые изменения морфологии, физиологии, поведения тихоокенских лососей, равно как своеобразие экологической обстановки на местах размножения и форма заботы о потомстве. Вскрыта адаптивная сущность некоторых из указанных преобразований и невыLLL GLLL LLLLLL G SLLLLLLLLLL aLLLLL LLLL LLLL LLLLLLLLS ной особи, так и в развитии вида. Анализ указанных изменений и особенностей биологии размножения с позиций теории этапности развития привел нас к убеждению в необходимости выделения нерестового этапа (Смирнов, 1959 б, 1964 б, 1965; Еремеева, Смирнов, 1965; Еремеева, 1967). Тихоокеанские лососи — рыбы моноцикличные и их онтогенез завершается нерестовым этапом. Этим они существенно отличаются от видов рода Salпо, рыб полицикличных. В результате сложных морфофункциональных преднерестовых преобразований у производителей изменяются требования к внешней среде, благодаря чему они выходят на соответствующие участки водоемов и мечут икру в такой обстановке, которая благоприятствует развитию потомства. Тем самым обеспечивается преемственная связь смежных поколений и продолжение рода. У разных видов и внутривидовых экологически отличных форм преднерестовые изменения и перест происходят в специфической среде. Знания этой специфики необходимы для разработки экологического метода стимуляции созревания производителей.
Переходя к сравнительному анализу особенностей развития, необходимо подчеркнуть следующее. Результаты морфо-экологического исследования тихоокеанских лососей и литературные данные убеждают в том, что каждому виду в различные периоды жизни, этапы и на отдельных стадиях наряду с общими чертами, свойственными всем представителям рода Опсоrhynchus и другим рыбам соответствующей экологической группы, присущи те или иные черты специализации. Внутри видов также имеются группировки, существенно различающиеся экологией размножения, что находит отражение и в морфогенезе. Яркий пример модификационной изменчивости производителей и хода развития их потомков представляет карликовая форма нерки (Смирнов, 1959 a, 1964 б). Последняя мельче анадромной, малоплодовита и персд нерестом изменяется слабо. Из мелких икринок карликовых особей зародыши вы
лупляются призначительно более низком уровне дифференцировки, а последующее развитие протекаст ускоренно. Другим примером глубокого изменения хода онтогенеза может служить сдвиг полового созревания у карликовых самцов симы, чавычи, кижуча на мальковый период жизни (являние неотении). В связи с созреванием в раннем возрасте и до миграции в море, без существенной смены среды, карликовые самцы нс прстерпевают значительных преднерестовых изменений и, что особенно примечательно, приобретают способность участвовать в размножении повторно, становятся полицикличными. Примеры пластичности процесса развития далеконе исчерпываются приведенными. Указав некоторые из них, мы хотели подчеркнуть широкий диапазон изменчивости. В дальнейшем сравнительном обзоренс предполагается рассматривать все разнообразие видовой и внутривидовой изменчивости морфогенеза лососей. Это слишком увеличило бы объем работы. Будут обсуждены лишь некоторые материалы, представленные в описательной
aCTH IICCIe OBaHHH.
Отличия раиних стадий морфогенеза и процесса эпиболии. При сравнcнии стадий дробления обнаруживаются различия размеров, формы и взаимного расположения бластомеров. У большинcтвa тихоокеанских лососей бластомеры относительно крупныс и в два слоя располагаются при образовании 16 клсток. У ксты тенденция к их двуслойному расположению иногда наблюдается даже на стадии 8 бластомеров (Дислер, 1957). Иная картина свойственна нерке. Ее бластомеры имеют сходный диаметр, но лежат свободнее и в два слоя располагаются после пятого деления, на стадии 32 бластомеров (см. рис. 21 Д, Е и рис. 32Д, 51, Е, 60Д). Сходная картина отмечена унcскольких представителей рода Salпо (Иванов, 1937; Стефанов, Денчева, 1964/1965; Вернидуб, 1967 а). Указаннос отличие можно объяснить меньшим количеством цитоплазмы, которая занимает, однако, сходную с другими видами площадь чем обеспечивается закладка достаточно крупного зародыша, свойCTBECHHOTO (COCHIM,
Различается темп морфогенеза. Например, дробление у нерки идет быстрее, нежели у чавычи (Combs, 1965). По данным М. Я. Иевлевой (1951), зародышевая полоска у кижуча и кеты появляется раньше, чем у горбуши и нерки. В дальнейшем эти расхождения увеличиваются. Брюшные плавники закладываются у ксты по сравнению с кижучом позже на 78 градусо-дней, а у нерки — на 198 градусо-дней. В описательной части содержится много материалов для аналогичных сопоставлений. Существенно различается время появления первичных зародышсвых клеток, закладки и дифференцировки гонад. Из лососей только горбушс свойственна протогиния. У этого вида гонады формируются особенно быстро, что стоит в связи с коротким циклом жизни (Персов, 1965, 1966, 1968).
Вариации количества желтка, цитоплазмы и их соотношения в яйцах лососей отражаются на продолжительности процесса эпиболии. Раньше других видов желточный мешок покрывается бластодермой у нерки, как правило, на стадии 23—26 сомитов. У кижуча процесс эпиболии завершается при 29—33 сомитах, в крупных яйцах чавычи, ксты, симы — лишь после образования 52—58 сомитов, у горбуши он иногда затягивается до формирования 63—64 сомитов. Соотвстственно указанным различиям, другие процессы формообразования протекают не в одинаковой ситуации. Так, кровообращение у нерки устанавливается много позже завершения эпиболии, тогда как у кеты, чавычи, симы и горбуши — непосредственно после исчезновения желточной пробки, а иногда и при ее наличии. С продолжительностью эпиболии связаны сроки сохранения икрой повышенной чувствительности к механическим воздействиям, поскольку устойчивость наступает после завершения этого процесса. Различается и уровснь чувствительности яиц к механичсским раздражениям. Сходное воздействие вызывает наибольшую гибель икринок нерки, несколько меньшую у симы, кижуча и самую низкую у икринок кеты и горбуши (Смирнов, 1954 б, 1955 б, б; Крыхтин, 1962, Иевлева, 1967). Более чувствительны к травмированию икринки с тонкой оболочкой.
Особенности развития кровеносной системы и эмбриональной моторики. В связи с развитием в среде, обедненной кислородом, и большой величиной икринок эмбрионально-личиночные органы дыхания у лососей достигают очень мощного развития. Дыхательную функцию у эмбрионов и личинок выполняют сосудисты с системы, покрывающие поверхность желточного мешка, развивающиеся в субэпидермальном слое покрововверхней поверхности головы и на туловище, мандибулярные нгиоидные дуги аорты, капилляры парных, непарных и хвостового плавников. Довольно мощного развития достигают псевдобранхии. Жаберные лепестки закладываются в конце инкубации икры. Жаберное дыхание устанавливается после вылупления; его роль возрастает уличинок и особенно у мальков после образования чешуйного покрова. Параллельно развитию и интенсификации функции жабер эмбрионально-личиночные органы дыхания редуцируются.
В развитии кровеносной системы наблюдаются видовые различия и модификации, связанные с особенностями режима инкубации икры. Например, у кижуча в Соколовском инкубаторе сердце начинало пульсировать на стадии 43—45 сомитов, а в Адотымовском — при 47-49 сомитах. У нерки, развивавшейся в речной воде, то же состояние наступало до образования 44 сомитов, а при инкубации включевой воде с низкой температурой — на стадии 44—46 сомитов. У осенней кеты, симы, перки кровообращение начинается, как правило, после образования 52—54сомитов, но в условиях хорошей аэрации может задерживаться на время, достаточное для формиРования ещc десяти сомитов. У горбуши, развивающейся в лучших кислородных условиях, ток крови появляется не раньше образования 58 сомитов. У тех лососей, которые развиваются в условиях меньшего насыщения воды кислородом, кровь раньше приобретает интенсивную окраску, быстрес формируются кардинальные вены, развивается печеночное кровообращение и подкишечно-желточная сосудистая система заменяется печеночно-желточной. Cсть капилляров на желточном мешке максимальной густоты достигает ко времени вылупления. Исключение составляет анадромная нерка. В ее икринках эмбрионы развиваются дольше, чем у других видов, и мелкие сосуды на желточпом мешке начинают редуцироваться до вылупления. В наших измерениях длина капилляров на 1 мм2 площади желточного мешка у горбуши достигала 6,5 мм, у осенней кеты и чавычи 7,5—8,0 мм, а у нерки 10 мм (ко времени вылупления нерки эта величина снижалась на 1,5—2,0 мм). УкаРис. 67. Сеть капнллярных сосудов занные различия иллюстрируна желточном мешке во время вы- ет рис. 67. Большое значение yo", 臀 #: для нормального развития лоучастки радиусом 1 мм c середины сосеи имеет проточность грунлевой стороны желточного мешка та. Ее отсутствие, как было описано на примере чавычи, может приводить к аномалиям в формировании кровеносной системы, наиболее значительным в каудальной области. В дальнейшем это ведет к замедлению роста и деформации хвостового отдела, рождению уродов или гибели эмбрионов.
Составной частью механизмов внешнего дыхания зародышей рыб являются протоплазматическая и нервно-мышечная моторика. Ее развитие у лососей описано достаточно подробно (Дислер, 1953, 1954, 1957, 1960: Wilker, 1953; Смирнов, 1958, 1960 a, 1964; Резниченко, 1959 a, б; Котляревская, 1968 а, б). Видовые различия моторики четко выражены в развитии функции грудных плавников. У большинства лососей они приобретают подвижность в течение девятого этапа развития (у кеты после 215 градусо-дней, у нерки —240, усимы и чавычи примерно после 340 градусо-дней), но у горбуши только в конце десятого этапа развития (при 370 градусо-днях). Примечательно, что с началом работы грудных плавников деятельность осевой мускулатуры на значительное врсмя ослабевает. Очевидно, работа грудных плавников обеспечивает достаточное перемешивание перивителлиновой жидкости, а в смысле расхода энергии она экономичнее, нежели движения довольно крупного эмбриона. Позже развивается подвижность челюстей и жаберных крышек. Функциональная активность осевой мускулатуры вновь возрастает перед вылуплением, когда эмбрионы под оболочкой начинают энергично вращаться. Как и само вылупление, этот моменту различных видов и форм приходится на разные стадии дифференцировки. Вылупившиеся зародыши на некоторое время вновь успокаиваются, тогда как частота двшжеIIий грудных плавников возрастает. Их энергичная работа обеспечивает постоянную смену воды у дыхательных поверхностей. С этого жс момента неуклонно возрастает роль жаберного дыхания. Вариабельность всличины икринок, толщины оболочек, мощности эмбриональных органов дыхания, как и условий аэрации гнезд, должна отразиться на интенсивности двигательной активности эмбрионов. К сожалению, сравнительных исследований в этом направлении на лососях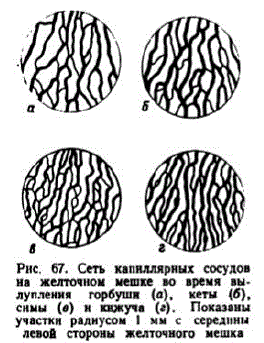
екоторые особенности пигментации зародышей. Обнаруживаются видовые различия сроков, порядка появления хроматофоров, их расположения и обилия. Раньше других видов хроматофоры появляются усимы. У нее первые липофоры на верхней поверхности головы были замечены вначале девятого этапа развития, при длине эмбрионов около 10 мм, а меланофоры-в начале десятого этапа. К этому времени длина эмбрионов увеличилась примерно па 3 мм. У кижуча липофоры имеланофоры на голове выявляются в начале десятого этапа, когда длина эмбрионов немного превышает 10 мм. У чавычи, как и усимы, первыми появляются липофоры, но позже, при длине эмбрионов 14—15 мм, меланофоры же становятся заметными при достижении эмбрионами длины 16 мм. У остальных видов раньше обнаруживаются меланофоры. У кеты и нерки это происходит в начале десятого этапа (при длине соответственно около 14 и 13 мм), а у горбуши — в середине того же этапа. Первые липофоры у нерки замечены в середине десятого этапа, укеты в его конце, у горбуши лишь в начале одиннадцатого этапа, когда эмбрионы достигали длины 15—16 мм. Слабее других видов пигментированы эмбрионы, личинки и мальки горбуши. Быстрее и обильнее пигментируются эмбрионы чавычи, кижуча, особенно же
У потомков одной и той же самки чавычи в условиях низкой температуры и хорошей аэрации раньше появлялись меланофоры (см. рис. 42, Б), а при более высокой температуре и изменчивом газовом режиме первыми и на более высоком уровне дифференцировки выявлялись липофоры (см. рис. 42, В). Сдвиги в развитии пигментации при разном режиме инкубации икры присущи и друTHM BHJLa M.
Продолжительность инкубации и уровень развития вылупляющихся зародышей. Перед тем как перейти к сопоставлению следует отметить трудность сравнения сроков инкубации лососей. Дело в том, что в зависимости от условий развития вылупление происходит в разном возрасте, после прохождения различного количества градусо-дней. Кроме того, у одного и того же вида выход из икринки смещается на разные стадии морфогенеза и даже на смежные этапы (Смирнов, 1959 a, 1964 а, б; Котляревская, 1968 а, б).
Сам процесс вылупления у одних видов проходит за относительно короткое время (сима, кста, чавыча), у других сильно растянут (горбуша, особенно нерка). Поэтому целям сравнения отвечают наблюдения, проведенные над икрой разных видов, развивавшейся в сходных условиях, в то же время отвечающих естественным требованиям каждоголосося. Сопоставление, проведенное с соблюдением указанных условий (Смирнов, 1964 б), показало, что массовое вылупление раньше началось у кижуча и симы (при 445 и 476 градусо-днях, соответственно), а дольше других видов инкубация икры продолжалась у горбуши и нерки (601 и 628 градусодней). В той же примерно последовательности возрастает и уровень развития зародышей ко времени массового вылупления. Кижуч и сима выходят из икры в начале дифференцировки лучей непарных плавников (см. рис. 54, Б, 62, Г.); у кеты и горбуши этот процесс заходит дальше (см. рис. 11, Г, 36, Б). Особенно высоким уровнем развития выдсляются зародыши, вылупляющиеся из икриноканадромной перки (см. рис. 26 А, Б). Из икринок же карликовых самок нерки эмбрионы выходят очень рано (см. рис. 26 Г). Кажется естественным предположение о большей продолжительности инкубации икры урано персстующих лососей и сокращении сроков инкубации икры у форм с поздним осснним и зимним нерестом. Примеры горбуши и, с другой стороны, кижуча отвечают такому представлению. Однако короткий срок инкубации икры летненерестующей симы с этой позиции понять нельзя. Очевидно, экологические зависимости сложнее, чтобы их понять, надо учесть особенности морфогенеза и режима нерестилищ. Складывается впечатление, что вылупление происходит раньше у тех лососей, у которых водоснабженис гнезд к концу инкубации по тем или иным причинам ухудшается, зародыши под оболочками на более ранних стадиях начинают испытывать недостаток в кислороде. Вылупление улучшает условия обмена и обеспечивает зародышам возможность переместиться в такие участки, которые омываются лучше. Наоборот, продолжительный срокинкубации икры мог выработаться в условиях более устойчивого режима водоснабжения. Это свойственно, как указывалось, нерестилищам проходной перки. Определенной стабильностью водоснабжения отличаются и русловые нерестилища горбуши (Никольский, 1952), а также камчатской чавычи; по продолжительности инкубации икры они стоят на втором и третьем местах вслед за неркой. Морфобиологические особенности молоди. Благодаря большому запасу питательных веществ зародыши в персстовых буграх развиваются долго, достигают значительных размеров, высокого уровня дифференцировки и способности энергично плавать. Такая молодь после выхода из гнезд успешнее избегает хищников. Кроме того, она способна питаться относительно крупными беспозвоночными. Пищей личинкам служат главным образом бентические организмы. Это связано со спецификой горных и предгорных рек, бедных IITAH KITOHOMI.
280
Как мы видели, все лососи могут переходить на внешнее питание еще при значительном остатке желтка. Этап смешанного питания у этих рыб охватывает целиком личиночный период жизни. Между тем в природе время приобретения зародышами способности захватывать и переваривать внешнюю пищу нередко наступает значитсльно раньше фактического перехода на активное питание. Причиной тому могут быть отсутствие корма или неблагоприятные внешние условия, такие, как низкая температура, слабая освещенность. Возникает вопрос о последствиях задержкиперехода лососей на активное питание, «относительного голода», как назвали такое состояние Кронфельд и Шеминский (Р. Ктоп[cld, F. Scheminsky, 1926). Указанные авторы наблюдали задержку росталичинок форели, если после резорбции желтка более чем наполовину они не получали внешней пищи. Было высказано предположение, что такие особн начинают испытывать недостаток каких-то веществ, необходимых для нормального развития. Очевидно, личинки должны обеспечиваться пищей своевременно. На сахалинских рыбоводных заводах ранее наблюдались случаи истощения молоди. После предварительных экспериментов была введена подкормка личинок кеты и горбуши; требование о проведении подкормки молоди этих видов включено в Инструкцию по искусственному разведению тихоокеанских лососей.
Отсутствие или плохое качество пищи нарушает развитие и ведет к быстрой гибели личинок многих рыб (Костомарова, 1962). Личинки лососей менее болезненно переживают недостаток пищи. Как было показано на благородном лососс и кете, это связано с тем, что при завершении рcзорбции желтка питательные вещества расходуются не полностью, а частично откладываются в полости тела личинок в виде жировой ткани (Hein, 1906; Дислер, 1954, 1957). Мы могли убедиться в том, что указанная особенность свойственна всем тихоокеанским лососям как, вероятно, и другим рыбам той же экологической группы. Резервные жировые отложения позволяют молоди пережить временную бескормицу, успешнее находить пищу в пресных водоемах или достигать приустьевых и морских участков, богатых кормом.
При сравнении молоди разных видов тихоокеанских лососей привлекает внимание укороченный пресноводный цикл жизни кеты и горбуши. Они в массе мигрируют в море, имся небольшой остаток желтка. Некоторое количество кеты задерживается в рсках более продолжительное время, но в пресных водах эти два вида не зимуют. Именно у них в отличие от остальных лососей отсутствуют карликовые самцы и жилые формы. Экологическая причина ранпего выхода в море достаточно определенна. Она состоит в том, что чрезвычайно многочисленное потомство горбуши и ксты в нсрестовых водоемах не находит достаточной пищи для продолжительного откорма. Молодь рассматриваемых видов ведет стайный образ жизни, что имеет защитное значение. Элиминирующее воздействие хищников в реках снижается, кроме того, благодаря
28
осуществлению катадромной миграции огромными массами и пренмущественно в ночнос врcмя,
От кеты горбуша отличается особснно ранним скатом. Ее молодь в подавляющей массе покидает-рски до начала активного питания, и не задерживается позже завершения этапа смешанного питания. Такая однородность сужает адаптивные возможности. В сравнении с другими лососями личинки и мальки горбуши менее высокотелы, имеют короткие плавники, слабее пигментированы и лишены пятен на боках тела (см. рис. 37, B—Д). Нектонный тип окраски можно объяснить тем, что молодь рано переходит к обиTHHKO BI TOT LILLER BOLEN.
Кета в реках живет немного дольше горбуши. Из коротких южных рек преобладающая масса ее молоди выходит вскоре после перехода на внешнее питание. В больших реках часть популяции проводит весь первый, а небольшое количество особей значительную часть второго малькового этапа развития. Момент перехода к катадромной миграции у кеты не фиксирован так строго, как у горбуши. В Амуре дольше задерживается молодь летней кеты (Солдатов, 1912). Личинкам и малькам этого вида свойственна пятнистая окраска, имеющая криптическое значение (см. рис. 14, Б—Г; 15). С возрастом молодь расселяется по реке и придаточным водоемам, благодаря чему-используются ббльшие кормоBE TOLLA LH.
Молодь нерки (см. рис. 28, Б, В, и верх рис. 30), как и у первых двух видов, постоянно держится в стаях. По экстерьеру и окраске ее личинки и мальки похожи на кету (четкие диагностические признаки указаны при описании видов). Среди тихоокеанских лососей нерка выделяется особснно продолжительной пресноводной фазой жизни. В нерестово-вырастных водоемах постоянно живут особи, находящиеся на разных этапах и периодах развития, что благоприятствует более полному использованию кормовых ресурсов. По сравнению с родственными видами нерка в наибольшей мере удовлетворяет свои пищевые потребности за счет пресных водоемов.
В озерах нерка питается преимущественно планктоном. Характеру пищи отвечает развитие длинных и многочисленных жаберных тычинок своеобразной формы (см. рис. 27). С переходом из литорали в пелагиаль молодь приобретает серебристую нектонную окраску. При жизни в мелководных ручьях и выращивании в прудах пятнистая окраска сохраняется продолжительное время. В реках и ручьях большой удельный вес в питании нерки занимают бентос и воздушные насекомые (Грибанов, 1948; Крогиус, 1951, Семко, 1954; Симонова, 1972 б). Из таких нагульных водоемов мальки могут мигрировать в озера, крупные лимнокрены и задерживаться в них. Однако значительное количество нерки с ручьевых и речных нерестилищ мигрирует в море сеголетками и даже личинками. Мелкая молодь в море подвержена более высо
282
кой элиминации, нежели крупная. Этим объясняется повышенная плодовитость особей, размножающихся в ручьях ирсках.
Молодь кижуча, чавычи и симы рано утрачивает тенденцию к образованию стай, рассредоточивается по водоемам, занимаст на мелководьях индивидуальные участки и охраняет их. Между разноразмерными мальками устанавливаются иерархические отпошения. В поведении и габитусе молоди этих видов много общих черт. С занятых позиций мальки совершают броски за проплывающими кормовыми объектами. Спектр питания молоди очcнь широкий. Значительный удельный вес в рационе занимают представители авифауны. Так как мальки используют разнообразную пищу и не образуют плотных скоплений, они продолжительное время могут жить в водоемах, относительно бедных водными кормовыми организмами. Соответственно обитанию у дна и своеобразному способу охоты мальки высокотелы, имеют длинные парные плавники, высокие спинной и анальный плавники с удлиненными передними ветвистыми лучами, крупную хвостовую лопасть. Преанальная складка у этих видов растет долго, достигает большой величины и продолжительное время сохраняется. Они выделяются также яркой пятнистой окраской. Большое количество молоди данной группы видов в пресных водах живет не менее года. Всем им свойственно явление неотении и повторное участие карликовых самцов в нересте. Усимы и кижуча имеются жилые формы.
Наряду с отмеченным сходством молодь каждого вида отличается строением, поведением, излюбленными местами обитания, что отражается на составе рaционов. У кижуча особенно долго сохраняется преанальная складка; это косвенно указывает на продолжительное обитание в придонных участках. Его молодь избирает тихие, часто сильно заиленныс протоки и старицы, прибрежья со свешивающейся в воду растительностью, заходит для нагула в озера, а мест с быстрым течением избегает. Рацион молоди кижуча отличается особенно высоким содержанием воздушных насекомых, которых он успешно отлавливает с поверхности, выпрыгивая из воды. Такому поведению отвечает форма тела и строспие плавников (см. рис. 55, B; 56). Сравнивая особенности экологии, окраски и миграционного поведения молоди тихоокеанских лососей, Ноат (1958) счел возможным поведение молоди кижуча принять за наиболее близкое к исходной речной форме, а образ жизни, характер окраски и миграционное поведенис молоди горбуши — наиболее специализированным. У рассмотренных видов мы видели четко выраженное соответствие конституции, окраски, образа жизни, в частности миграционного поведения, занимаемой ими экологической ниши. То же характерно и для других видов.
По внешнему виду и поведению мальки чавычи напоминают кижуча. Отличаются они несколько меньшей высотой непарных плавников, относительно слабой их пигментацией, особенно анального плавника, иным расположением хроматофоров (см. рис. 45
283
Г. 46). Мальки чавычи могут удивительно быстро менять окраску соответственно фону окружающего грунта и их очень трудно обнаружить среди прибрежных камней, где они скрываются (Edmunson et al., 1968; Reimers, 1968; собственные наблюдения). Интересно, что молодь осенненерестующей чавычив отличие от летней, в масcе мигрирует в море сеголетками, даже до закладки чешуи (Rutter, 1904; Mason. 1965).
Молодь симы особенно высокотела и выделяется своим внешним видом (см. рис. 65, Г, Д; 66). Личинки питаются преимущественно бентосом и некоторое время живут стайками. Мальки занимают индивидуальные участки, пищу добывают подобно кижучу, но не выпрыгивают из воды за насекомыми. С возрастом пищевой спектр расширяется. У симы особенно ярко выражена эврифагия. Главная особенность рациона состоит в изобилии разнообразных беспозвоночных, вырастающих вне водоемов. Поэтому мальки и карликовые особисимы и могут жить в горных реках и небольших притоках с очень бедной фауной. Сима реофильнее кижуча, раньше выходит из грунта и использует прибрежные мелководья, ее молодь держится в более верхних участках рек.
При анализе межвидовых и внутривидовых отношений внимание исследователей привлекает распространение среди молоди лососей хищничества. Действительно, в желудках годовиков и двухгодовиков кижуча, симы, чавычи, нерки, гольцов и других видов иногда в значительном количестве встречается мелкая молодь лососей, в том числе того же вида. В каждом случае важно проанализировать причины хищничества. При этом рассматриваются особенности режима водоемов, их населения, обеспеченность рыбы кормами, условия обловов, причины концентрации молоди и потенциальных хищников. В противном случае могут складываться ошибочные представления о масштабах хищничества и каннибализма, что отражается на рыбохозяйственных рекомендациях, определении путей наилучшего использования нерестовых фондов, выборе объектов разведения, содержании работ по биотической мелиорации нерестово-вырастных водоемов. Внимательный анализ биоценотических отношений показывает, что в природе не так часто наблюдаются условия, приводящие к скоплению разновозрастной молоди одного или нескольких видов. Однако подобная ситуация нередко складывается у плотин, заграждений для учета покатников, у мест выпуска молоди из питомников и т. п., что благоприятствует развитию хищничества и каннибализма (Hunteг, 1959; Rickсг, 1962: Леванидов, 1969; наши наблюдения). Исследования питания и распределення разновозрастной молоди в нескольких водоемах Сахалина недали оснований говорить о существенном воздействии крупных мальков симы и кижуча на численность других лососей (Крыхтин, 1955, 1962; Калмыкова, 1957; Воловик, 1963, 1964; наши наблюдения). В обследованных камчатских озерах кижуч поедает значительное количество рыбы, но это молодь девятииглой и трехиглой колюшек, корюшки, являющихся трофически
284
ми конкурентами нерки (Зорбиди, 1970а). Точно так же и гольцы рода Salveliпus не могут относиться к бесспорно вредным для лососевого хозяйства объектам. В некоторых водоемах они потребляют мало рыбы или же истребляют пищевых конкурентов молоди нерки и приносят этим скорее пользу, чем вред (Крогиус, Крохин, 1948, 1956; Савваитова, 1961, 1973).
Выше говорилось о видовых и внутривидовых различиях производителей и мест размножения, а также адаптивных особенностях эмбриогенеза. Молоди также присущи черты морфофункциональной и экологической специализации, своеобразие поведения. В естественных условиях личинки и мальки каждого лосося занимают в той или иной мере отличающуюся стацию и пищевую пишу, которые, как и вся система адаптаций, меняются с возрастом, при переходе с этапа на этап. Это следует внимательно учитывать при анализе биоценотических отношений и разработке схем рациональной эксплуатации бассейнов нерестовых водоемов. Важно иметь в виду, что видовая и популяционная разнокачествен
LLG S S LLLLL S LLLLLLaLLLLLL S LLLL S LaLLaLLLL S aLLLLLLLLLL возможности нерестово-вырастных водосмов и кормовые ресурсы морей и океана.
Смотрите также
Бассейновый метод выращивания лососевых рыб