
Развитие симы
Половые продукты. В садках самцы Pಖ್ಖ овали сперму до 19 сут., давали 8—9 порций объемом от 1,4 до 52 мл (среднее 12,8 мл) и общий объем спермы достигал 81 мл. Количество сперматозоидов в 1 мм3 колебалось от 16,8 до 36,8 млн. (в среднем 21,23 млн.). В эякуляте на 1 г живого веса самцов число сперматозоидов достигало 307 млн., т. е. почти в 5 раз большей величины, нежели у горбуши.
При 9,3° через 20 сек. после разведения спермы водой ею воплодотворялось 90% икринок, через 1 мин. — 60%, а через 2,5 мин. оплодотворялись единичные икринки (рис. 59). Тот же график показывает быструю утрату в воде фертильности и икринKEMH
Зрелые икринки весят от 126 до 226 мг (М=174, 79+3,22, о = 25,2). Четырехлетние самки чаще давали икру крупнее, чем трехлетние. Икра имеет оранжево-розовый или красно-оранжевый цвет с малиновым оттенком (варнации цвета икры лежат в пределах длины волны 592—594 нм). Состав икры приведен в табл. 1. В 1959 г. дважды одновременно оплодотворялась икра симы и горбуши и эти виды скрещивались. Икра развивалась в Соколовском инкубаторе, режим которого описан выше. Синхронность осеменения и одинаковые условия развития материала придают ему большую сравнительную ценность. При описании икру, оплодотворенную 10/VIII 1959 г., будем называть первой серией, а оплодотворенную 13/VIII — второй. В 1954 г. развитие симы изучалось на Адотымовском заводе. Сведения о развитии симы даются в работе М. Л. Крыхтина (1962) и наше описание будет кратким.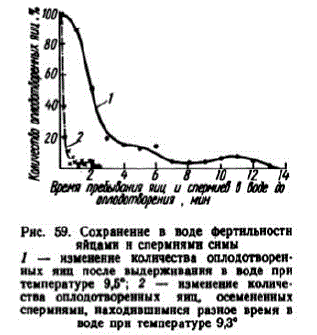
Рис. 59. Сохраненне в воде фертильностн яйцаминспермнями снмы I - изменение колнчества оплодотворенных яиц после выдеринвания в воде прH температуре 9,5"; 2 — изменение количества оплодотворенных яиц, осемененных спермнямн, находившнынся разное время в воде прн температуре 93"
Эмбриональный, или зародышевый, период развития
1. Обводнение осемененных икринок, образование зародышевого диска. Процесс обводнения икринок при 9,2° длился 45— 50 мин. Икринки разных самок во время набухания прибавляли в весе от 6,3 до 15% (в среднем на 11,5%). Диаметр набухших икринок варьирует от 6,1 до 7,5 мм, чаще составляет 6,8 мм. Диаметр некоторых икринок достигает 8 мм. Толщина оболочек икринок, начавших дробиться, колеблется от 32,4 до 50,1 мк (М=39,6 +0,49). Яйцевые оболочки прозрачнее, нежели у кеты и горбуши. У осемененной икры интенсивность дыхания возрастает. При 14° 1 кг икры за час потребляет 52—55 мг кислорода. Затем эта величина снижается до 10—12 мг/час, сохраняясь примерно на таком уровне до начала кровообращения 1.
При температуре 8,4° зародышевый бугорок сформировался
1. Здесь н далее сведения о потребленннннрой кислорода н темпе развитня симы на Краснопольском заводе приводятся по М. Л. Крыктину (1962).
Данные об измененнн чувствнтельности нкры к механическим воздействиям взяты у того же авторану Хата (Hata, 1ძრწწ.
через 12 час. после осеменения. Его диаметр равнялся 0,9—1,1 мм. высота — 0,45—0,50 мм.
2 этап. Дробление зародышевого диска. При той же температуре дробление начиналось через 13—14 час. и требовался еще 1 час, чтобы два бластомера сформировались у всех яиц. Второе
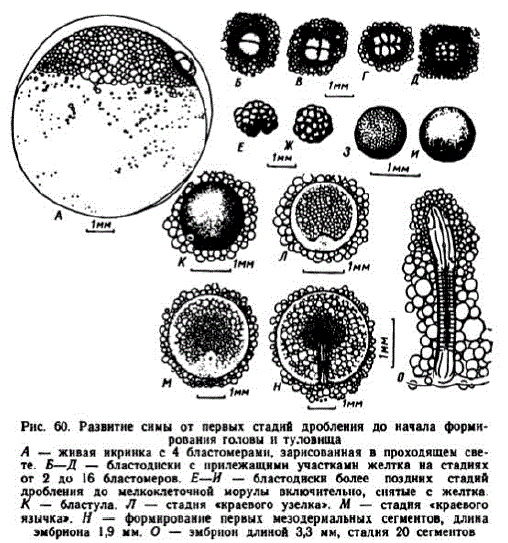
Рис. 60. Развитие симы от первых стадий дробления до начала формирования TOTUEL II туловнща А — живая нкринка с 4 бластомерами, зарисованная в проходящем свете. Б-Д - бластоднски с прилежащими участкамн желтка на стадиях от 2 до 16 бластомеров. Е—И — бластоднcкн более поздннх стадий P ления до мелкоклеточной морулы включительно, снятые с желтка — бластула. Л. — стадня «краевого узелка». М — стадия «краевого язычка». Н — формнрование первых мезодермальных сегментов, длина эмбрнона 1,9 мм. О — эмбрион длиной 3,3 мм, стадия 20 сегментов
четвертое дробления наступали с периодичностью около 4 час. После третьего деления контур зародышсвого диска принимал форму овала с малым диаметром 0,73—0,98 мм и большим — 0,82—1,15 мм (рис. 60, A—Г). Четвертая борозда дробления проходила в экваториальной плоскости. Через 30 час. насчитывалось 28—32 бластом сра, а чсрез 34—35 час. число бластомеров в икринках, развивавшихся быстрес, достигало 64 (рис. 60, Ж). К исходу вторых суток диаметр поверхностных бластомеров уменьшился до 80—70 мк, а в возрасте 3 суток — до 30 мк.
3 этап. Бластула. За несколько часов до исхода пятых суток зародышевые диски начинали увеличиваться; к их окончанию (42,5 градусо-дней) диаметр бластодисков достигал 1,3—1,6 мм, формировалась бластула (рис. 60 К). В возрасте 5 сут. 6 час. бластодиски имели диаметр 1,6—1,8 мм. Клетки поверхностного слоя уменьшились до 19—20 мк, а лежащие под ними — до 22— 23 мк. Этап длился немного более суток.
4 этап. Гаструляция. В 6-суточном возрасте (51,5 градусодня) диаметр бластодисков составлял 2—2,3 мм. Они стали плоскими, на их поверхности располагались клетки, имевшие диаметр 16—18 мк. Началась дифференцировка зародышевых пластов (рис. 60, Л). К исходу 8 сут. диаметр эмбрионального щита достиг 2,5—3 мм. Образовался «краевой язычок» длиной 0,8—1,2 мм (рис. 60, M). К исходу 10-х суток диаметр эмбрионального щита достиг 3,5 мм. При средней температуре 8,7° этап длился около 4 сут.; у зародышей с замедленным темпом развития он продолжался на полсуток дольше. При температуре 10—16" этап завершался за 2—3 сут.
5 этап. Формирование головы и туловища зародыша. В возрасте 10 сут. 2 час. (86 градусо-дней) бластодиск имел диаметр 3,4—4 мм и покрывал примерно третью часть желточного мешка. Зародыш вырос до 2—2,2 мм. Появились первые сомиты, начали дифференцироваться отделы головного мозга, наметились зачатки глаз (рис. 60, H). На 11-е сут. при 11—12 сомитах были видны зачатки слуховых пузырьков, начали формироваться хрусталики.
В возрасте 12 сут. бластодерма охватывала примерно половину сферы желточного мешка. У зародышей длиной 3,3 мм насчитывалось до 20 сомнтов. Стали заметными обонятельные плакоды (рис. 60, О). У эмбрионов длиной 4,2 мм насчитывалось до 30 сомнтов и обособилась хвостовая почка. После того как бластодерма закрыла больше половины желточного мешка, чувствительность икры к механическим агентам заметно понизилась. В Соколов
ском инкубаторе этап продолжался около 6 сут., в Краснопольском - 4 сут.
6 этап. Обособление задней части тела зародыша от поверхности желточного мешка. 16-суточные эмбрионы (150 градусодней) имели длину 4,6—5 мм и у них насчитывалось 43—44 сомита. Появились хвостовые миотомы и задний участок туловища начал обособляться от желточного мешка. На этой стадии, то есть несколько раньше, нежели у кеты, чавычи и горбуши, стенки сердечной трубки начали волнообразно сокращаться (рис. 61,A). К началу этапа сформировались хрусталики, появились мелкие отолиты, выявились зачатки жаберных крышек. Бластодерма покрыла почти весь желточный мешок. В течение этапа заверпается процесс эпиболии. Желточная пробка закрывалась в середине или в конце 17 сут. (155—160 градусо-дней) при 52—53 сомитах, а чаще — при 54—56 сомитах. В отдельных случаях эпиболия затягивалась до 18-суточного возраста и образования 59 сомитов. Как и у других лососей, чувствительность икры в это время высокая.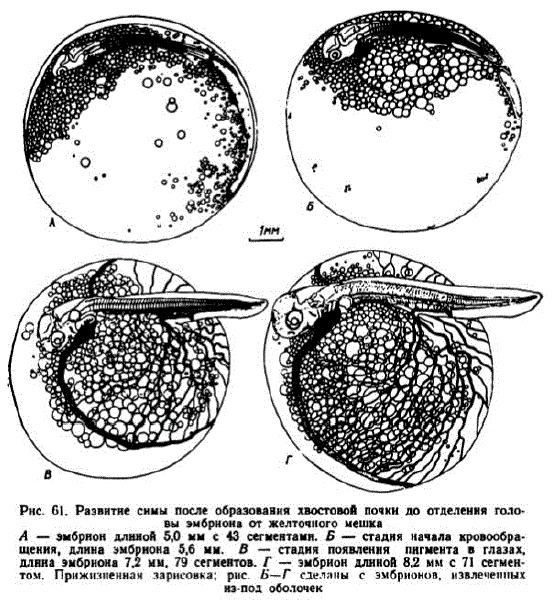
Pнс, 61. Развитне снмы после образования хвостовой почки до отделення голоB эмбриона OT KelTORIOTO MelIKa A — эмбрион длиной 50 мм с 43 сегментами. Б — стадия начала кровообращения, длина эмбрнона 5,6 мм. В - стадня появлення пнгмента в глазах, длнна эмбриона 7,2 мм, 79 сегментов. Г - эмбрион длиной 8,2 мм с 71 сегменTOM, Прижизненнан зарнсовка; рис. эмбрионов, Belle
нз-подоболочек
7 этап. Развитие подкишечно-желточиой системы кровообращения. Циркуляция крови отмечалась в возрасте 17 сут. у зародышей длиной 5,2—5,6 мм, имевших от 52 до 55 миотомов. У некоторых из них в это время еще сохранялась небольшая желточная пробка (рис. 61, Б). Эмбрионы, у которых кровообращение начиналось позже, имели не более 59 миотомов и длину 5,8 мм. Первоначально кровь по спинной аорте протекала только до 1—2-го хвостовых миотомов и содержала мало форменных элементов.
В начале этапа намечается ротовая воронка, выявляются зачатки нижних челюстей, голова начинает отделяться от желточного мешка. Позади зачатков грудных плавничков живых эмбрионов просматриваются клубочки пронефроса. На том же уровне с правой стороны выявляется зачаток печени. Эмбрионы энергично поводят хвостом и изредка переваливают тело то на один, то на другой бок.
М. Л. Крыхтин (1962) отметил начало кровообращения усимы после формирования 65 сегментов и достижения эмбрионами длины 7,8—8,3 мм. Его сообщение о движении крови в это время почти до последних сегментов хвоста свидетельствует об описании не начальной стадии кровообращения, а более поздней.
Вскоре после начала кровообращения сосуды обильно заполняются эритроцитами, длина хвостовых артерий и вены увеличивается, на желточном мешке появляется все больше тонких капилляров. После начала кровообращения существенно возрастает интенсивность обмена. При температуре воды 14" и содержании кислорода 10,6 мг/л в течение 1 час. 1 кг икры потреблял 15 мг кислорода. В Соколовском инкубаторе при 9,2° этап длился около 12 час.
8 этап. Возникновение кардинальных вен и смешаниого подкишечно-желточного и печеночно-желточного кровообращения. Кардинальные вены образовались в возрасте 18—19 сут., когда эмбрионы достигли длины 5,5—6 мм. У таких зародышей по хвостовой артерии кровь проникала более чем на 20 миотомов. Из хвостовой вены часть ее текла в задние кардинальные вены, остальная — в подкишечную вену. Имелись правая и левая ветви краевой вены желточного мешка. На нем заметно возросла площадь, покрытая капиллярами, и их густота. Возникло печеночное кровообращение.
В течение этапа абсолютная длина головы увеличивалась мало, а относительная — немного уменьшилась. Наблюдался ускоренный абсолютный и относительный рост хвостового отдела, разрастались непарные плавниковые складки. У отдельных эмбрионов в возрасте 21 сут. (197 градусо-дней) в глазах появился
К исходу 22 сут. эмбрионы достигли длины 6,8—7,2 мм (рис. 61, B). Голова отделилась от желточного мешка. Длина хвоста составляла до 30% общей длины и в нем насчитывалось 30— 32 миотома. Основання грудных плавников приняли наклонное положение. Появилась задняя брыжеечная артерия.
К концу этапа закончилась сегментация хвоста. Глаза стали серыми. В статоакустических органах четко выявились полукружные каналы и образовались крупные отолиты. Капиллярами покрылось более половины поверхности желточного мешка. Сформировалось две пары жаберных артерий. По всему туловищу образовались сегментальные сосуды (рис. 61, Г). При температуре 9,5— 9,7° этап продолжался около 7 сут.
9 этап. Формирование печеночно-желточной системы кровообращения. В возрасте 25—26 сут. (235—245 градусо-дней) подкишечно-желточная вена запустевала и с этогб времени жслточный мешок снабжался кровью только через печень. Эмбрионы имели длину 8—8,5 мм. К указанному времени спинная плавниковая складка разрослась до первых миотомов и в дальнейшем только расширялась. Наметилась закладка непарных плавников. У эмбрионов длиной около 12 мм (развитие с 13/VIII по 14/IX 1959 г.) голова стала высокой, но ее относительная длина сократилась до 17,3% общей длины. Глаза почернели. Жаберные крышки закрыли первую пару жабер. Хвостовой отдел превысил 37% всей длины, при этом число его миотомов с 32—34, имевшихся на предшествующем этапе, сократилось до 28—29. Сформировались 3 гипуралии, заложились первые лучив лопасти плавника, начала развиваться капиллярная сеть, хвостовой плавник утратил симметрию. Грудные плавники заметно удлинились (pHc. 62, A).
На 28-е сутки началось кровоснабжение третьей, а через сутки и 4-й пары жабер. Появились гноидные дуги аорты. Увеличилась сеть капилляров на верхней поверхности головы. Сегментальные сосуды развились в большей части хвостовых миотомов. Капиллярами покрылся весь желточный мешок. Желточная вена приняла вентральное положение. В возрасте 29 сут., т. е. немного раньше, нежсли угорбуши и кеты, на верхней поверхности головы появились многочисленные мелкие липофоры яркой красной окраски. При температуре 9,6—9,9° этап длился 4,5—6 сут.
10 этап. Дифференцировка верхних и нижних конусов миотомов. У некоторых эмбрионов в возрасте 36 сут. (343 градусо-дня) в хвосте выявились зачатки верхних и нижних конусов миотомов (рис. 62, Б). Зародыши достигали длины 12,7—14 мм. У них обозначились зачатки верхних челюстей. На 2-й и 3-й жаберных дужках появились зачатки жаберных лепестков. Стали заметными псевдобранхии. Число хвостовых сегментов сократилось до 25—26. Заложились брюшные плавнички. Грудные плавники приобрели подвижность. Сегментальные сосуды образовали сеть капилляров над спинным мозгом и у поверхности тела. На 1 мм2 левой стороны желточного мешка длина капилляров достигла 6,7—7,0 мм. В этом возрасте, то есть значительно позже липофоров, на голове появились первые меланофоры. Выявились железки вылупления. Эмбрионы приобрели способность хвостом производить волнообразные движения. Скороткими интервалами они ритмично машут грудными плавничками. Возросла интенсивность дыхания. За 1 час при температуре 10-11“ 1 кг икринок потреблял до l7— 18,8 MT KHcJIopoJa.
На Соколовском заводе при средней температуре 10° этап длился 8—9 сут.: на Адотымовском заводе к этому времени температура понизилась до 4,2° и этап продолжался более полумссяца. 11 этап. Развитие подвижности челюстей, жаберных крышек,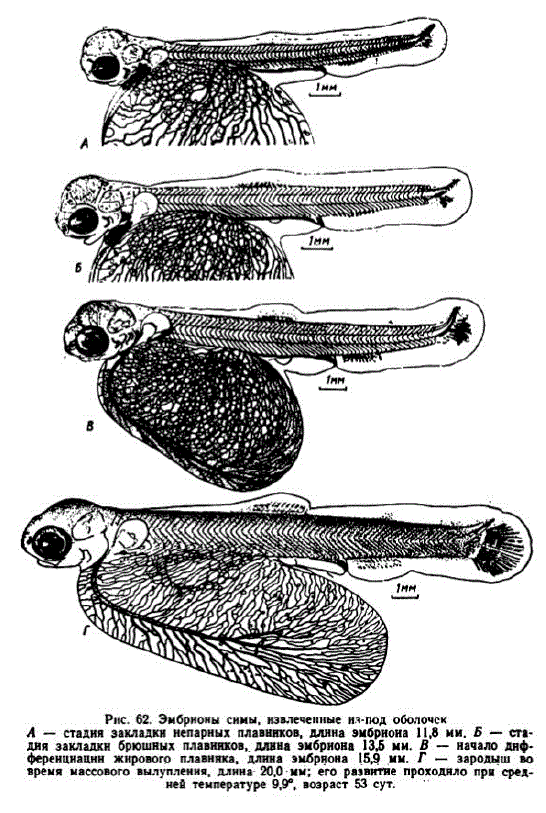
завершение инкубации. К началу последнего этапа инкубации икры эмбрионы увеличились примерно до 15 мм. В Соколовском инкубаторе этап начинался на 38—40-е сут. К этому времени увеличившиеся нижние челюсти полностью прикрыли ротовое отверстие. Челюстями и жаберными крышками эмбрионы совершали энергичные движения. Слуховые пузырьки максимально сблизились с глазами. Продолжал дифференцироваться хвостовой плавник. В спинном и анальном плавниках закладывались опорные элементы. Пигментация тела усилилась; появились меланофоры неврального и гемального рядов.
К 42-м сут. (405 градусо-дней) эмбрионы достигли длины 15,5—16 мм. На Адотымовском заводе, где осенью температура резко понижается, аналогичное состояние отмечено в возрасте 70 сут. (рис. 62, B).
ри температуре 10,3—10,7° этап длился 10—14 сут., а при средней температуре 4,3°— 24—27 сут.
12 этап. Пассивное состояние свободных зародышей. В первой серии за время инкубации температура изменялась в пределах 8,4—10,7°; первые эмбрионы вышли из икринок на 48 сутки, массовое вылупление проходило на 52—53-е сут. (518—528 градусо-дней), а последние эмбрионы вылупились на 56-е сут. (560 градусо-дней). В икре других серий наиболее ранним было вылупление в возрасте 器 сут. (452 градусо-дня). Сима вылупляется дружно. Раньше выходили зародыши из мелкой икры. На Адотымовском заводе, где температура колебалась в пределах 8,3—4,5°, вылупление началось на 68-е сут. (423 градусо-дня) и было массовым в возрасте 80—83 сут. (до 494 градусо-дней). На Краснопольском заводе икраинкубировалась 35—48 суток (450— 550 ¥:
ри постоянной температуре 16,2° инкубация икры жилой симы длилась 28,5 сут., а при 5,8°—84 сут. Наибольший выход нормальных эмбриоиов наблюдался при температуре от 8 до 11"; в этих условиях инкубация завершалась за 505—508 градусо-дней (Kawajігі, 1927).
В мелкой икре эмбрионы достигают длины 16—18 мм, в крупных икринках — 20—21 мм. Особираннего срока вылупления имеют в хвосте 11—13 лепндотрихий, расчленение которых начинается, а в период массового вылупления насчитывается до 17—20 лучей и в средних лучах имеется по три членика.
В нашей первой серии массовое вылупление проходило при длине от 18,7 до 203 мм (рис. 62, Г). Эмбрионы имели 37—39 туловищных и 23—25 хвостовых миотомов. Хвостовой отдел, измеренный от переднего выступа первого хвостового миотома, достигал 39% длины зародыша, а при измерении от анального отверстия составлял 35—36% всей длины. Из непарной плавниковой складки вычленились анальный, спинной плавники и обозначился контур жирового плавника. В грудных плавниках начали формироваться лучи.
Ко времени вылупления глаза зародышей приобрели подвижность. Жаберные крышки полностью прикрывали жабры. В жабрах и псевдобранхиях развивалась сеть капилляров. Почти завершилось образование перегородок обонятельных отверстий. Органы боковой линии за инкубацию достигли такого же уровня развития, как и у кеты (рис. 63,A). Имелся небольшой зачаток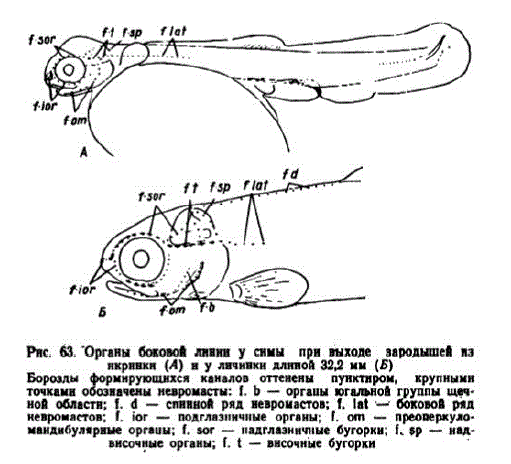
Рис. 63. Органы боковой линии у симы при выходе зародышей из нкрннки (A) ну личннки длиной 32,2 мм (Б)
Борозды рмирующихся каналов оттенены пунктиром, крупными
точкамн обозначены невромасты: f. b — оргашы югальной группы щеч
ной области; 1. d — спинной ряд невромастов; f. lat - боковой ряд
невромастов: I. іог — подглазничные органы; I. оп — преоперкуломанднбулярные оргашы, І. ѕог — надглазннчные бугоркн; І. 5p — над
височные органы; f.t — внсочные бугорки
плавательного пузыря, наметилось вычленение желудка. На ". вой жабре заложилось 3—5 жаберных тычинок (рис. 64, A). По уровню развития сима при вылуплении сходна с кижучем. Среди всех лососей ее зародыши выделяются интенсивной пигментацией. Как и у других лососей, вылупившиеся зародыши ведут себя пассивно них реакции на внешние раздражители нечетки. В малоактивном состоянин при температуре 10,6° зародыши симы находятся 8—11 сут., а при 3,8°— 18—25 сут.
13 этап. Формирование непарных и брюшных плавников, развитие плавательного пузыря. В возрасте 60—64 сут., когда сима достигала длины 23—25 мм, она стала перемещаться навстречу потоку, положительно реагировать на прикосновение посторонних предметев, отрицательно относиться к свету.
Рот зародышей принял полуконечное положение (рис. 65, A). В кожистых перепонках жаберных крышек появились лучи. Лучи хорошо выявились в спинном, анальном, грудных плавниках и начали формироваться в брюшных плавниках. Густота сосудистой сети желточного мешка сократилась, уменьшились его объем и площадь, не закрытая миотомами.
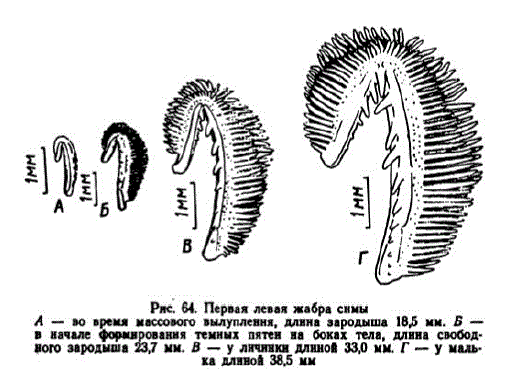
Рис. 64. Первая левая жабраснмы A — во время массового вылуплення, длина зародыша 18,5 мм. Б — в начале формировання темных пятен на боках тела, длина свободного зародыша 23,7 мм. В — уличннки длиной 330 мм. Г - у малька длиной 38,5 мм
Зародыши приобрели темную окраску, причем много хроматофоров появилось на непарной плавниковой складке. В отличие от родственных видов, у симы хроматофоры имеются и на преанальной плавниковой складке вблизи ануса. В коже наряду с красными липофорами появились более мелкие хроматоформы оранжевого цвета.
При температуре 9,4—9,6° через 25—30 сут. после вылупления на боках тела появляются темные пятна (рис. 65, Б). К этому времени на челюстях прорезаются зубы. Хвост принял симметричную форму и в нем наметилась выемка. Его лучи начали разветвляться, заложилось несколько редуцированных лучей. Брюшные плавники приобрели подвижность. Преанальная складка продолжала растн. Тело свободных зародышей стало более обтекаемым. Возросла вязкость желтка и жировые капли перестали в нем перемещаться. Появились первые пилорические придатки, начали формироваться петли кишечной трубки, наблюдалась елабая перистальтика. Зачаток плавательного пузыря достит длины 3—3,5 мм; начался его быстрый рост. В питомниках такие зародыши время от времени поднимаются в толщу воды, но долго
зародыши время от времени поднимаются в толщу воды, но долго
плавать они еще не могут и вскоре опускаются на дно.
В Соколовском питомнике при температуре 9,2° этап длился
25—30 сут., в Адотымовском при 2,7°— 2,5—3 месяца.
Смотрите также
Бассейновый метод выращивания лососевых рыб