
Эмбриональный, или зародышевый, период развития кижуча
1 этап. Обводнение икры, образование зародышевого диска. Примерно через 10 мин. осемсненные икринки приобретают слабую клейкость, которая исчезает после смыва водой. При TE2= ратуре 6° процессоводнения икры длится 70—75 мин. (рис. 50).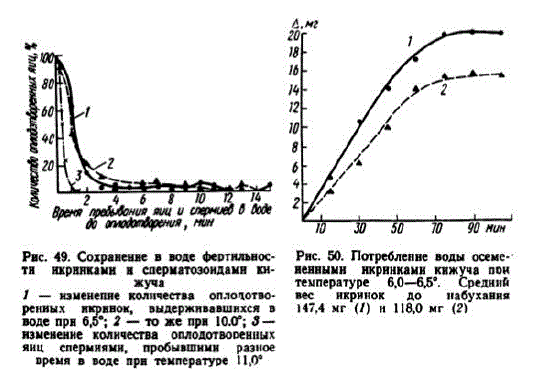
яиц спермняын, пробывшнын разное время в воде при температуре 110"
В наших опытах икра разных самок в результате обводнения прибавляла в весе от 12,6 до 15,8% (средняя прибавка веса у икры 7 самок составила 13,5%). Мелкие икринки после набухания имели диаметр 5,8 мм, крупные — 7,3—7,5 мм. Толщина оболочек у набухших икринок колеблется между 36 и 57 мк (M=473+0,55). Поверхность набухших икринок становится белесоватой, матовой.
Как и у других видов, до попадания в воду икрипки стойко переносят механические воздействия. С началом обводнения чувствительность к толчкам, вибрациям резко возрастает и снижается через короткое время после набухания (табл. 15). Во время на
Таблнца 15

Примечанне. Икра инкубировалась при температуре 9,6 -8". Икринкина разных стадиях подвергались воздействию вибрации в течение 210 сек.
бухания у анимального полюса икринок собирается большинство жировых капель, начинается концентрация цитоплазмы. Сформировавшийся зародышевый диск имеет диаметр от 1,05 до 1,2 мм и высоту 0,4—0,5 мм.
2 этап. Дробление зародышевого диска. На Соколовском заводе у икры, оплодотворенной 19/X 1953 г. (первая серия) и развивавшейся вначале при 9,5°, дробление началось через 8 час. 50 мин. после осеменепия. В возрасте 12 час. большинство икринок имело 4 бластомера, в 18 час. — 8 и к завершению первых суток от 12 до 28 бластомеров. В полуторасуточном возрасте их число превышало 100. Через 2 сут. образовалась морула, поверхностные клетки которой имели диаметр 30—45 мк. Икра второй серии была оплодотворена 22/X 1954 г. и инкубировалась на Адотымовском заводе. В начале развития среднесуточная температура равнялась 4,6°. Ход изменения температуры показан на рис. 5. У этих икрипок дробление началось в возрасте 20 час. и к исходу вторых суток образовалось 64 бластомера.
После первого деления основание зародышевого диска принимает овальную форму, длинная ось этого овала достигает 1,3—
1,4 мм, а на стадии 8 бластомеров — 1,55 мм. После 4-го и 5-го делений основание зародышевого диска становится круглым, бластомеры плотно прилегают друг к другу, диаметр бластодиска уменьшастся. Дробящийся зародышевый диск немного погружается в желточную массу (рис. 51, А, Б). С началом дробления
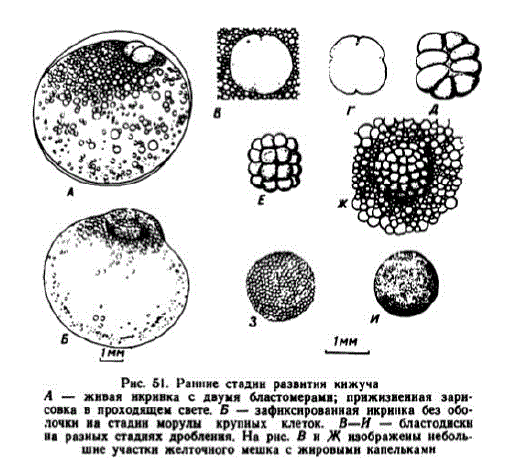
Pнс. 51. Ранние стадин развитникижуча А — живая икрннка с двумя бластомерами; прижизненная зарисовка в проходящем свете. Б — зафиксированная икринка без оболочки на стадин морулы крупных клеток. В-И — бластодискн на разных стаднях дроблення. Нарнс. В и Ж изображены небольLHe yqacTKH KeHTONHOTO NeuKa C жнровыми HELIJAKANH
чувствительность икринок к вибрации повышается. Икра кижуча к такому воздействию чувствительнее икры кеты и горбуши.
В конце этапа дробления при температуре 3,2—3,5" за час 1000 икринок потребляла кислорода 0,139 мг (в пересчете на 1 кг икры — 0,9 мг кислорода).
8 этап. Бластула. При температуре 9,6° стадия бластулы наступает к концу 3-х сут. Бластодиск к этому времени стал плоским и немного разросся. В возрасте 3 сут. 4 час. его диаметр достиг 1,6—1,7 мм и он полностью закрыл зону мелких жировых капелек (рис. 52, A). Поверхность бластулы состояла из плотно расположенных полигональных клеток диаметром около 25—30 мк. Ниже их рыхло располагались более крупные клетки. У икринок второй серии стадия бластулы наступала в возрасте 6 сут. и этап длился около 2 сут.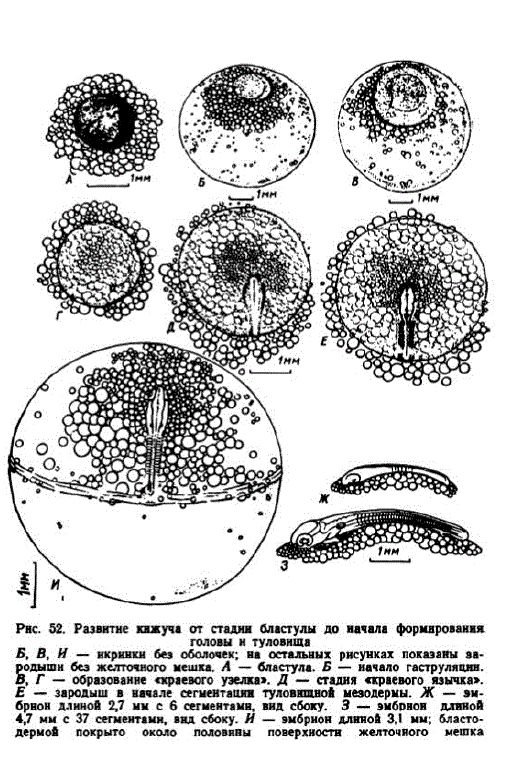
4 этап. Гаструляция. В первой серии к возрасту 4 сут. диаметр бластодисков увеличился до 2,2—2,4 мм, они начали закрывать более крупные жировые капли (рис. 52, Б). В возрасте 6 сут. (57,5 градусо-дней), когда диаметр бластодисков достиг 3—3,3 мм, образовался «краевой узелок» (рис. 52, В, Г), вскоре превратившийся в «краевой язычок» (рис. 52, Д.). Во второй серии гаструляция началась на исходе 8-х сут. (38,8 градусо-дней). В 14 сут. эмбриональный щит имел диаметр 5—5,3 мм и «краевой язычок» увеличился до 2 мм. С началом гаструляции чувствительность икры существенно возрастала.
5 этап. Формирование головы и туловища. В первой серии сегментация туловищной мезодермы началась в конце 8-х сут. (72—76 градусо-дней), во второй — в конце 16 — начале 17 сут. Эмбрионы с 3—4 сегментами имели длину 2,2—2,6 мм. У них образовались первичные мозговые пузыри. Диаметр эмбрионального щита к этому времени достиг 4 мм (рис. 52, E). У эмбрионов с 5—7 сегментами образовались зачатки глазных бокалов (рис. 52. Ж). Вскоре появились слуховые пузырьки, отстоявшие от глаз на расстоянии примерно равном ширине 10 сегментов (рис. 52, И).
Рост зародыша сопровождался быстрым обрастанием желточного мешка бластодермой. При средней температуре 4,3° на 20-е сут. бластодерма закрыла примерно половину сферы желточпого мешка, у эмбрионов образовалось 13—14 сегментов. Снизу и немного левее заднего конца хорды был виден купферов пузыPCK.
В сравниваемых сериях в возрасте 11 сут. (115 градусо-дней) и на 25-е сут. (105 градусо-дней) процесс эпиболии завершался. Близкая к этой стадия показана на рис. 53, А. В икринках, имевших диаметр 7,1 мм, к этому времени зародыши достигали длины 4,5—4,7 мм и имели 30—33 сегмента. В икринках диаметром 6,5 мм процесс эпиболии завершался немного раньше, при 29—31 сегменте и длине эмбрионов 4,3—4,5 мм. Это происходило раньше, чем у других лососей, исключая нерку. В сходные сроки обособляется хвостовая почка, которая при 37 сегментах достигает длины 0,7 мм.
В конце этапа при температуре 4,8° 1000 икринок за 1 час потребляли кислорода 0,63 и 1,07 мг (1 кг икры соответственно 3,79 и 5,26 мг). Крупная икра потребляет кислорода больше (вторые из "??????: цифр).
6 этап. Обособление задней части туловища зародышей от поверхности желточного мешка. Этап в условиях Соколовского инкубатора начинался в возрасте 12—12,5 сут. (115—120 градусодней), когда эмбрионы достигли длины 4,6—4,8 мм и имели 37—40 сегментов. Сегментация туловищного отдела к этому времени закончилась, он включал 38—40 ссгментов. Сформнровался задний отдел кишечника и анальное отверстис. Наметилась дифференцировка желудка. В аппаратах Адотымовского инкубатора
выявились энцефаломеры. Наметились зачатки жаберных крышек. Образовалась перикардиальная полость, в которой был виден зачатоксердца.
В течение этапа слуховые пузырьки увеличиваются, в них появляются скопления мелких отолитов. Длина головного отдела уменьшается, а высота головы увеличивается. Быстро парастают длина и число сегментов хвостового отдела, развивается непарная плавниковая складка. Задняя часть туловищного отдела постепенно обособляется от желточного мешка. Одновременно растет зачаток преанальной плавниковой складки. Эмбрион приподнимается Ha. KeITOIHLIM 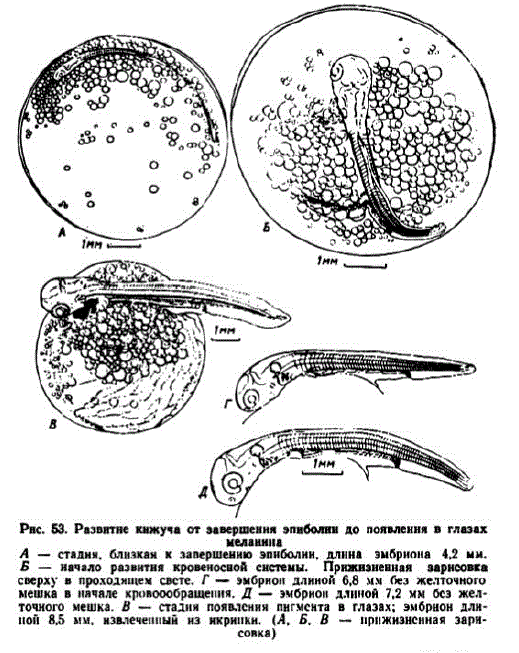
У некоторых эмбрионов в Соколовском инкубаторе сердечная мышца пачинала сокращаться при 39—41 сегментах, чаще при 43—45 сегментах, когда становились заметными сокращения туловинцных миотомов (14 сут., 134 градусо-дня). При более низкой температуре, в Адотымовском инкубаторе, становление нервномышечной моторики задерживалось до формирования 47—49 миотомов, когда эмбрионы достигали длины 6—6,3 мм (35-38 суток, около 145—155 градусо-дней). При температуре 3,2—3,5° 1000 таких икринок за час потребляет 0,556 мг кислорода (в пересчете Ha l Kr H KpHHoK — 2,5 Mr).
7 этап. Развитие подкишечно-желточной снcтемы кровообращения. Появление тока крови у разных эмбрионов первой серии отмечалось в 15 и 16 сут. (144—153 градусо-дня), во второй — на 38—40-е сут. (155—162 градусо-дня). Такие эмбрионы чаще имели длину 6,5—6,8 мм и у них насчитывалось 59—61 миотом (у некоторых 63), из которых 39—40 туловищных (рис. 53, Б, В). В Адотымовском инкубаторе, снабжаемом речной хорошо аэрированной водой с низкой температурой, кровообращение начиналось на несколько более поздней стадии дифференцировки.
К началу этапа абсолютная длина головы немного увеличилась, но ее относительная величина уменьшилась и составляла менее 25% общей длины эмбрионов. Высота головы продолжала увеличиваться. Она начала отделяться от желточного мешка. Выявились зачатки грудных плавничков. На живых объектах через вентральные концы 4—7-го миотомов просматривались первые клубочки пронефроса. Непарная плавниковая складка на спине разрослась до первых сегментов.
В начале этапа кровью снабжаются туловищные сегменты и лишь один-два первых хвостовых сегмента. Сосуды быстро и обильно заполняются форменными элементами. Вскоре увеличиваются хвостовые артерия и вена, подкишечно-желточная вена, на желточном мешке появляются тонкие капилляры. Постепенно возрастает подвижность эмбрионов.
8 этап. Возникновение кардинальных вен и смешанного подкншечно-желточного и печеночно-желточного кровообращения. В Соколовском инкубаторе менее чем через сутки, а в Адотымовском через 1,5—2 сут. после пачала кровообращения кровью снаб
жаются уже 10-12 хвостовых миотомов. Образуются задние кардинальные вены, кювьеровы протоки, появляются первые сосуды печеночно-желточной вены. В начале этапа эмбрионы имели длину 6,8—7,7 мм; в первой серии насчитывалось 68 миотомов, во второй — 68—70.
В течение этапа начинают функционировать первая и вторая пара жаберных артерий. Количество капилляров желточного мешка быстро нарастает. В описываемых сериях на 21—22-е сут. (200—210 градусо-дней) и 44-46-е сут. (175—182 градусо-дня) зародыши удлинились до 8,2—8,6 мм и в их глазах появился меланин (рис. 53, B). В Соколовском инкубаторе пигмент в глазах кижуча появлялся на сутки раньше, чем у горбуши, причем эмбрионы были немного крупнее. К концу этапа сегментация хвостовой мезодермы заканчивалась.
9 этап. Формирование печеночно-желточной системы кровообращения. Кровоснабжение желточного мешка из подкишечной вены прекратилось у эмбрионов длиной 8,6—9 мм; это происходило в первой серии в возрасте 23—24 сут. (218—227 градусодней), во второй серии — на 52—56-е сут. (202—217 градусодней). У эмбрионов насчитывалось соответственно 67—69 и 75—77 сегментов, из них туловищных было 37—38 и 42—43 сегмента. Голова у извлеченных из-под оболочек зародышей распрямлялась. Заметно увеличилась ее высота. Глаза стали крупными. Образовались овальные обонятельные ямки. Стато-акустические органы располагались от глаз на расстоянии, равном примерно ширине двух миотомов. В них ??????? полукружные каналы, отолиты стали крупными. Образовалась небольшая ротовая впадина. Зачатки жаберных крышек частично прикрыли первую пару жабер. Кончик хорды начал изгибаться кверху. Плавниковая складкана спине разрослась до первого мнотома. Заметно увеличилась преанальная складка, передний конец которой располагается на каудальной поверхности желточного мешка. Основания выросших грудных плавников приняли наклонное положение (рис. 53, B).
В начале этапа кровь течет по трем нарам жаберных артерий. На верхней поверхности головы появились капилляры. Кровью не снабжаются лишь 3—5 последних сегментов. Во всех туловищных и первых 10 хвостовых мнотомах имелись сегментальные артерии и вены. Сформировалась задняя мезентериальная артерия, капилляры которой охватывали задний участок кншки, средняя часть кишки была покрыта капиллярами передней мезентериальной артерин. В сосудистую ссть желточного мешка кровь поступала только из крупных печеночных вен. На протяжении этапа эмбрионы большее время оставались неподвижными, изредка наклоняли тело направый или левый бок и плавно изгибали хвост то вправо, то влево, кончиком хвоста доставая головы. К концу этапа, песмотря на понижение температуры до 2,7°, интенсивность дыхания повысилась, и в теченне 1 часа 1000 икринок потребляла кислорода 0,71 мг, а 1 кг икринок — 3,26 мг.
10 этап. Дифференцировка верхних и нижних конусов мнотомов. Начало формирования верхних и нижних конусов миотомов отмечено на 31—33 сут. (289—306 градусо-дней) и на 67—71-е сут. (254—267 градусо-дней). Эмбрионы к этому времени выросли до 10—11,3 мм. Разросшаяся нижняя челюсть немного выступала вперед вертикали, опущенной от зрачка. Глаза потемнели, в пих появились кристаллики гуанина. Слуховые пузырьки располагались непосредственно у глаз и длина головы относительно укоротилась. В полостях головного мозга появился розоватый диффузный пигмент. Увеличившиеся жаберные крышки прикрывали две пары жабер. Последние хвостовые миотомы стали распадатъся и число сегментов сокращаться. Дальнейшее удлинение хвоста проходило за счет увеличения ширины миотомов. Начали формироваться гипуралии, заложились первые лучи хвостового плавника, между ними появились капилляры. В местах закладки анального и спинного плавников плавниковая складка расширилась, образовались мускульные почки. Обнаружились признаки окостенения элементов плечевого пояса. Основания выросших грудных плавников приняли почти вертикальное положение; плавники стали снабжаться кровью, приобрели подвижность. С этого времени эмбрионы стали двнгаться редко.
В течение этапа появляется ток крови в 4-й паре жаберных дужек и в гиоидных дугах. На верхней поверхности головы развивается густая сеть капилляров. Сегментальные артерии и вены формируются во всех миотомах. В передней части туловища от сегментальных сосудов отходят капилляры к спинному мозгу и поверхности тела. Количество капилляров на желточном мешке продолжает увеличиваться. Полость кишечной трубки приобретает желтовато-зеленоватую окраску. На голове и сверху туловища появляются меланофоры и более мелкие вишнево-красные липофоры.
У эмбрионов первой серии в возрасте 35 сут. (324 градусодня), а во второй — на 74-е сут. (216 градусо-дня) заложились брюшные плавнички. В это же время выявились зачатки верхних челюстей. В обонятельных ямках начали формироваться перегородки. После витальной окраски метнленововой синью можно было различатъ зачатки жаберных лепестков. Обозначилось небольшое число железок вылупления. С усложнением формы миотомов эмбрионы приобрели способность совершать волнообразные движения хвостом и туловищем.
11 этап. Развитие подвижности челюстей, жаберных крышек и завершенне инкубации. В Соколовском инкубаторе 11 этап начался в возрасте 40—43 сут. (372—397 градусо-дней), в Адотымовском — в возрасте 86—91 сут. (309—321 градусо-день). Этап завершается вылуплением. В его начале эмбрионы имели длину 14—15,5 мм. У извлеченных из оболочек эмбрионов желточный мешок принимал яйцевидную форму. Его задний конец отвисал. Диаметр некоторых жировых капель увеличился до 1—1,5 мм.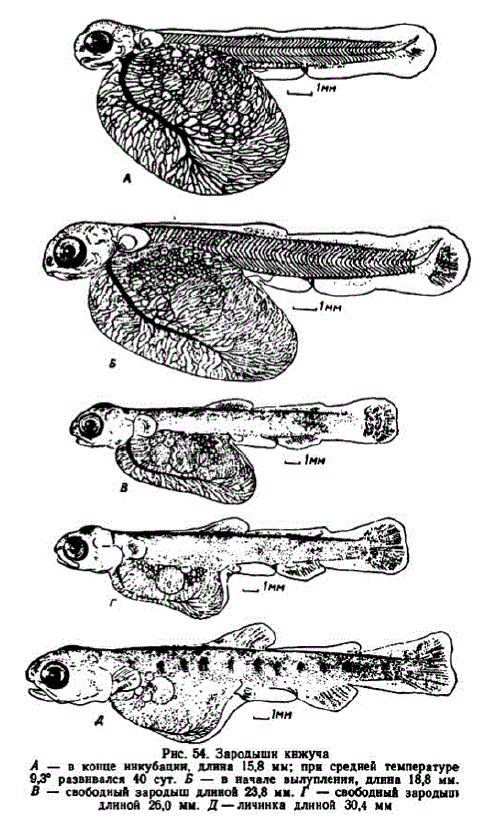
Из произошедших изменений отметим прежде всего относительное сокращение длины головы, которая стала составлять примерно 17% длины эмбриона. Жаберные крышки закрыли жабры. Край нижней челюсти достиг вертикали, опущенной от переднего края глаза. В течение этапа прорывается ротовая воронка. К концу этапа жаберно-челюстной аппарат приобретает слабую подвижность. Глаза черные, иридируют; их диаметр более 1,2 мм.
В результате резорбции эмбриональной плавниковой складки четче определились контуры анального и спинного плавников, в основаниях которых образовались крупные мускульные почки. Обозначился зачаток жирового плавничка. Стали хорошо заметными брюшные плавнички. В хвосте заложились до 5—6 лепидотрихий. В хвостовом отделе начали формироваться позвонки.
Густота сосудистой сети на желточном мешке продолжала нарастать. На рис. 54, А хорошо видна мощная нижняя желточная вена и впадающая в нее правая ветвь краевой желточной вены, уже заметно редуцировавшейся.
Усилилась пигментация дорзальной поверхности головы и тела. Единичные меланофоры появились в спинном и хвостовом плавниках, а также над спинным мозгом. Возросла и плотность расположения липофоров. Кроме дорзальной поверхности они распространились в непарную плавниковую складку и хвостовой плавпик. Хорошо видны железки вылупления.
До времени, близкого к вылуплению, эмбриопы под оболочкой лежат спокойно и лишь редко наклоняют тело вправо или влево, взмахивая при этом хвостом, или производят хвостом волнообразные движения. Постоянное перемешивание перивителлиновой жидкости обеспечивается ритмичными движениями грудных плавничков. Выполнению той же задачи способствуют движения рта и мощная пульсация сердца, движения которого вызывают колебания массы желтка. Перед вылуплением эмбрионы возбуждаются и начинают подоболочкой энергично вращаться.
12 этап. Пассивное состояние свободных зародышей. Из икры, оплодотворенной в разные сроки с 14/X по 23/X 1953 г. и развивавшейся в Соколовском инкубаторе, вылупление начиналось в возрасте от 50 до 54 сут. (445—486 градусо-дней) и продолжалось 3—5 дней. В Адотымовском инкубаторе, где к этому времени вода охлаждалась до 2,5—2,2°, вылупление растягивалось почти на месяц. Сроки инкубации кижуча в разных условиях показаны в табл. 16.
Судя по имеющимся данным, кижуч развивается при меньшем диапазоне температуры, нежели родственные виды. При средней температуре 11,4° отмечено повышение отхода икры (Allen, 1958). По нашим наблюдениям, в начале инкубации крат. Ковременное воздействие температуры 12—13,5° не вызывало наРУшения развития и увеличення отхода. О нижнем пределе ожно судить по данным экспериментальных и заводских инкубаторов. В Паратунском инкубаторе развитие начиналось при
3,7—4° и заканчивалось при 2,3°. На Ушковском заводе эмбриогенез проходит при устойчивой температуре около 4,5—4,1°. На Адотымовском — за инкубацию икры вода остывала с 5—4,5° до 2,5—2°. В Карымайском инкубаторе кижуч развивался при изменении температуры от 3,5 до 0,8°. Р. С. Семко (1954) называет этот вид наиболее холодолюбивым.
Таблица 16
На рис. 54, Б изображен эмбрион в начале массового вылупления. Размер зародышей связан с величиной икринок. Из икринок диаметром 6,3 мм они выходили при средней длине 20,2 мм, наиболее крупные достигали 21 мм. В мелкой икре до вылупления кижуч достигал 16—19 мм. После вылупления желточный мешок принимает яйцевидную форму. Его задний конец почти достигает начала хвоста и немного опущен. Срок инкубации кижуча относительно короткий и за это время желтка расходуется немного. На Соколовском заводе за инкубацию вес желтка уменьшался примерно на 13,5%. У вылупившихся зародышей имеется обычно одна крупная жировая капля, диаметром 2—2,4 мм, окруженная MeU KHIMH Ka Teb Ka MH.
Голова ко времени вылупления становится высокой, но составляет лишь 16—17% длины зародышей. Слуховые пузырьки максимально сближаются с глазами, диаметр которых достигает 1,4 мм. Выявились 3—4 зачатка жаберных тычинок. Жаберные крышки, в кожистой оторочке которых появились лучи, прикрыли жабры с хорошо развитыми лепестками. Постанальный отдел заметно удлиннлся и составлял более 38% общей длины зародышей. Число миотомов в хвосте сократилось до 25—27. В хвостовой лопасти развились гипуралии и 15—17 лепидотрихий (при раннем вылупленин насчитывалось по 10 лепидотрихий, а у поздно вылуп
212
ляющихся эмбрионов их число увеличивалось до 20). Ко времени вылупления зачатки лепидотрихий появились в спинном и анальном плавниках. Слабым выступом контура плавниковой складки, скоплением пигментных клеток и мезенхимы обозначился зачаток жирового плавника. Лопасти грудных плавников стали крупными, больше ширины 5 туловищных миотомов. Изменения сосудистой системы сводятся к следующему. Появились капиллярные сосуды в лепестках жабр и псевдобранхий. Капилляры оплели глазодвигательные мышцы. Развилась обильная сеть капилляров на дорзальной поверхности головы. Сосуды, отходящие от сегментальных артерий, образовали разветвления над спинным мозгом и в покровах тела зародышей. Увеличилась сеть капилляров между лучами хвостового плавника. К вылуплению на поверхности желточного мешка сеть капилляров стала очень густой, длина капилляровна 1 мм? достигала 9,5 мм и более.
Ко времени вылупления заметнее обозначилось вычленение желудка. Поперечная складчатость внутренней стенки кишки была выражена слабо. Зачаток плавательного пузыря имел вид небольшого пальцеобразного выступа, длиной около 1,5 мм.
При выходе из оболочки кижуч интенсивно пигментирован. Меланофоры покрыли переднюю и верхнюю поверхность головы, отдельные клетки появились на жаберных крышках. На теле меланофоры распространились до нижних конусов миотомов. Они имелись в непарных плавниках и в основании непарной плавниковой складки позади спинного плавника. Хорошо выявились периневральный, пернваскулярный и перитониальный пигментные ряды. На поверхности головы н тела между меланофорами располагались мелкие вишнево-красные липофоры. В непарной плавниковой складке и в хвосте липофоры заняли большую площадь, нежели меланофоры, но в анальном плавнике они еще не IORIBHTHCb.
Как и у других лососей, зародыши после вылупления успокаиваются. Глаза их еще неподвижны (подвижность появляется в конце этапа). Движения жаберно-челюстного аппарата слабее. Частые ритмичные движения грудных плавников создают ток воды вдоль тела и желточного мешка. Реакции на внешние воздействия аналогичны описанным у кеты. Сходно и состояние органов чувств системы боковой линии.
В лаборатории во время вылупления И. С. Васильевым было проведепо сравнение интенсивности дыхания зародышей, находившихся в икринках и только что вылупившихся. Определения проводились в сосуде с циркулирующей водой при температуре 5—5,5°. Тысяча икринок за 1 час потребляли кислорода от 45,7 до 76,7 мг (в среднем 56,5 мг), а вышедшие из оболочек зародыши — от 81,2 до 114,5 мг (в среднем 101,8 мг).
Особи, выходившие из икринок последними, достигали длины 21—23 мм. Среди задерживавшихся под оболочкой были и мелкие эмбрионы, имевшие длину 17 и даже 16 мм. У них обнаружены
2S
разные дефекты: уменьшенные глаза, слабо развитые непарные плавниковые складки, недоразвитые жаберные крышки, ослабленная сосудистая сеть. Сердце таких зародышей пульсировало медленнее. У некоторых из них были слабо развитыми железки вылупления.
13 этап. Формирование непарных, брюшных плавинков и плавательного пузыря. При температуре 4—6° через неделю после вылупления поведение зародышей менялось. Они начали реагировать на ток воды и двигаться ему навстречу, возбуждаться при освещении, положительно реагировать на прикосновение к посторонним предметам. При слабом течении в темноте зародыши лежали спокойно. Возбужденные, они всплывали в толщу воды, но плавали с трудом и короткое время. Видимо, в буграх с этого времени они могут начать рассредоточиваться, постепенно перемещаясь к поверхности.
К началу этапа рот принял полуконечное положение (рис. 54, B). Кожистый край жаберных крышек разросся до оснований грудных плавников. Непарная плавниковая складка впереди спинного плавника редуцировалась. Пpeанальная складка разрослась почти до заднего края желточного мешка. В лопасти хвостового плавника наметилась вырезка; в его лучах образовалось до 5 члеников. Хорошо выявились лепидотрихии анального, спинного и грудных плавников. Густота сети капилляров на желточном мешке заметно сократилась. Зародыши потемнели. Увеличилось и количество липофоров, особенно сверху головы и на спине. Липофоры появились в анальном плавнике, в основании преанальной складки, на грудных и брюшных плавниках.
При увеличении длины до 24,5—26 мм анальный и спинной плавники становятся высокими и обособляются (рис. 54, Г). В хвостовом плавнике появляются так называемые редуцированные лучи. Закладываются первые лучив брюшных плавниках. На челюстях появились зубы. В обонятельных ямках образовались перегородки. Продолжают разрастаться кожистые складки жаберных крышек. В отличие от других видов, преанальная складка у кижуча не останавливает свой рост и достигает нижней стороны желточного мешка. Желточная масса становилась вязкой и жировые капли перестали в ней перемещаться, когда зародыши меняли свое положение. В полости тела замечено отложение жира.
В течение этапа пропорции тела меняются мало. К концу этапа на боках тела появились темные пятнышки (рис. 54, Д). Желточный мешок к этому времени становится маленьким, приобретает обтекаемую форму, его вес уменьшается до 55—70 мг. Одновременно увеличиваются запасы внутриполостного жира. Впереди жирового плавника сохранился небольшой остаток непарной плавниковой складки. Хвост стал симметричным, его лучи ветвистыми. Вершины брюшных плавников достигли края преанальной складки. Последняя продолжала увеличиваться и в длину и в ширину. Напротив брюшных плавников ее ширина достигала или даже пре
214
высила 1 мм, а передний край располагался к голове ближе спинного плавника. Такой величины преанальная складка достигает только у кижуча.
В этом возрасте на церато- и эпибранхиалиях жаберных дужек насчитывалось до 10—12 тычинок. Крупные жаберные лепесткиразветвились. Продолжал формироваться желудок, внутренняя стенка кишки приобрела хорошо выраженную складчатость, наблюдались перистальтические движения. Быстро растущий зачаток плавательного пузыря у кижучат длиной 25 мм достигал 4,2—4,5 M M.
Свободные зародыши периодически всплывали к поверхности и совершали хватательные движения. В аквариуме они начинали захватывать мелкие планктонные организмы и другую пищу. Через 2—3 дня пищевая активность возросла и появились особи с хорошо заполненными кишечниками. В питомнике Адотымского завода при средней температуре 3,1° кижуч к плаванию в толще воды переходил через 33—38 сут. после массового вылупления. При температуре ниже 3,5" активность личинок была слабой.
. В. Крогиус при вскрытиях гнезд в Даниловском ключе (бассейн р. Паратунки) фиксировались пробы икры и личинок, которые были любезно предоставлены нам для просмотра. Судя по этому материалу, пятна на теле отдельных экземпляров кижуча появляются в конце февраля. Интересно, что в отличие от зародышей, выведенных в инкубаторах, у особей из гнезд желточный мешок не отвисал, и личинки имели более обтекаемую
форму.
Смотрите также
Бассейновый метод выращивания лососевых рыб