
Эмбриональный, нили зародышевый, период развития горбуши
В 1952 и 1953 гг. были проведены наблюдения за развитием икры горбуши, одновременно оплодотворенной икры осенней кеты, и полученными между этими видами гибридами (Смирнов, 1953а, 1954а, б, 1964а, б). В 1959 г. дважды удалось одновременно осеменить икру горбуши и симы и получить между ними помеси. Икра инкубировалась в одном рыбоводном аппарате, что позволило сопоставить темп развития указанных видов и их гибридов. Наблюдения проводились на Соколовском заводе; сведения о режиме его инкубатора приведены на стр. 38. При описании привлекаются некоторые наблюдения и над другими сериями икры, в том числе развивавшейся на Лесном заводе. Его инкубатор снабжается хорошо аэрированной речной водой и имеет режим, близкий к режимур. Лесной, охарактеризованному выше.
I этап. Обводненне осемененных икринок, этап образования зародышевого диска. Наблюдения за изменением веса обводняющихся икринок проведены при температуре 10,2°. Икру брали от двух самок. Икринки первой самки до помещения в воду имели средний вес 154,2 мг, второй — 127,3 мг. Интенсивнос обводнение проходило между 10 и 30-й мин., а закончилось через 40—50 мин. после помещения икры в воду. Набухшие икринки весили 166,5 и 138,1 мг; прибавление веса составило 8,0 и 8,5%. Набухшие икринки не строго шаровидны. Нам встречались икринки диаметром от 5,6 до 7,9 мм. Для развивающейся икры сахалинской горбуши указываются вариации в пределах 5,5—8,3 мм, при M= 6,9+0,004 (Галкина, 1965). Диаметр икринок амурской горбуши — 6,0 мм (Васильев, 1959б).
В процессе обводнения оболочка икринок становится прочной и теряет прозрачность. Ее поверхность покрывается белесоватым матовым налетом. Микропилярное поле, расположенное вблизи анимального полюса икринки и имеющее диаметр 1,6—1,8 мм, лишено белесоватого налета. Здесь расположено воронкообразное углубление, ведущее к микропилярному отверстию.
У развивающихся икринок, зафиксированных изотоническим раствором формалина, толщина оболочек варьирует от 55 до 80 мк (М=699+0,60). Оболочки икринок горбуши толще, нежели у других лососей и живая икринка выдерживает давление до 8—9 кг (Никольский, Соин, 1954).
Специальные опыты выявили существенное изменение чувствительности набухших икринок к механическим воздействиям (Смирнов, 1954б). Схема опытов аналогична той, которая применялась при изученни чувствительности икры кеты. В первой серии каждая очередная порция развивающихся икринок испытывала воздействие вибратора, пока его ось совершала 100 оборотов, во второй серии — 300 оборотов. Подопытная и контрольная икра инкубировались в одном аппарате при температуре 7,8—9,6°. Результаты воздействия определяли по проценту погибших икринок (табл. 9). Зрелые икринки до попадания в воду стойко переносят
Таблнца 9
 Начало формирования жаберных лепест
Начало формирования жаберных лепестKOB . . . . . . . O), 5 OO 53 Перед вылуплением O, O. OO
толчки. Вскоре после начала набухания чувствительность икры резко повышается, достигает максимума примерно через 30 мин., а затем постепенно снижается и примерно через 2 час. икра при
обретает устойчивость. Указанное время икра должна находиться
E OTOM TOKOL
2 этап. Дробление зародышевого диска. К началу дробления зародышевый диск принимает форму, близкую к полусфере, несколько суженной у основания. Его диаметр в яйцах горбуши из р. Мы составлял 0,7—0,9 мм (Васильев, 1959б), в яйцах горбуши из р. Калининки — 0,9—1,0 мм (Фроленко, 1959). Данные моих промеров бластодиска до стадии 4 бластомеров близки к данным И. С. Васильева. После 3 и 4-го делений бластомеры занимают немного большую площадь. Как и у кеты, бластодиск расположен над скоплением мелких жировых капель (рис. 32).
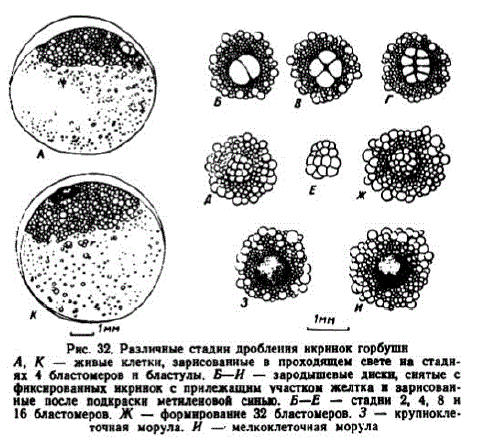
Рис. 32. Различные стаднн дробленнянкрннок горбушн А, К — живые клетки, зарнсованные в проходящем свете на стаднях 4 бластомеров н бластулы. Б—И — зародышевые диски, снятые с фикснрованных нкрннокс прилежащим участком желтка и зарисованные после подкраскн метиленовой синыo. Б-Е — стадни 2, 4, 8 и 16 бластомеров. Ж — формированне 32 бластомеров. З — крупнокле
точная морула. И — мелкоклеточная морула
При температуре 90-9,5 первая борозда дробления появлялась через 11—12 час. после осеменения, второе дробление наступало между 17 и 18 час., третье — через 21 час., четвертое — 26 час. В возрасте 30 час. образовалось 32 бластомера и в середине вторых суток — 64. Яйца дробились не синхронно; у некоторых из них наступление очередной стадии задерживалось на 2—2,5 час. На Лесном заводе при температуре 13,0—14,7° первое дробление происходило через 8,5 час. после осеменения, к исходу первых суток формировалось до 128 бластомеров.
1На ранних стадиях дробления зародышевые диски имеют высоту 0,4—0,5 мм. До стадии 4-х бластомеров зародышевые диски округлы. При следующем дроблении они несколько вытягиваются; овальная форма диска сохраняется и на стадии 16 бластомеров, но затем он опять принимает округлую форму. При четвертом дроблении борозды проходят в экваториальной плоскости и образуется два слоя клеток. В середине вторых суток развития образовалась морула крупных клеток, а к их концу — мелкоклеточная морула (рис. 32, И). В этом возрасте диаметр поверхностных бластомеров уменьшается до 60—65 мк.
Чувствительность икры к механическим воздействиям с началом дробления увеличивается. Во второй серии опытов отход такой икры после воздействия на нее вибрацией повышался до 20%, а с началом второго дробления — до 32%. Этого, однако, не было заметно при кратковременном воздействии вибратора.
3 этап. Бластула. При температуре 8,4—8,6° к завершению третьих суток зародышевые диски отдельных яиц начали разрастаться и их диаметр увеличился до 1,2—1,3 мм. При колебании температуры от 12,6 до 14,7° аналогичное состояние наступало через 42 час. Клетки верхнего слоя бластулы имели диаметр 30—35 мк, но у некоторых яиц уже уменьшились до 25—23 мк. Разрастание бластодиска служит указанием на формирование бластомерной бластулы. Со временем поверхностные бластомеры располагаются плотнее и образуют слой микроскопических полигональных клеток, отличающихся от рыхло расположенных нижних более крупных клеток. К концу этапа диаметр бластодиска достигал 1,8—2 мм и закрыл зону мелких жировых клеток (рис. 33, A). Чувствительность икринок к механическим воздействиям продолжала повышаться.
4 этап. Образование зародышевых пластов. К исходу 7-х суток (около 60 градусо-дней) диаметр бластодиска достигал 2 мм. Диаметр клеток эктобласта к этому времени уменьшался до 10—13 мк. Диск в центральной части становился настолько тонким, что через него просматривались расположенные ниже мелкие жировые капли. Края бластодиска покрывали часть крупных жировых капель; диаметр последних продолжал увеличиваться. В каудальной стороне зародышевого диска сформировался «зародышевый узелок» (рис. 33, Б). С разрастанием эмбрионального щита образовался «зародышевый язычок». Началась дифференцировка зародышевых пластов и хорды. В возрасте 9 сут. диаметр зародышевого диска был равен 2,5—3,0 мм, а длина зародыша 1,2—1,3 мм (рис. 33, В). На Лесном заводе при колебании температуры в пределах 11,0—14,7° гаструляция начиналась в возрасте 3,5 сут.; к исходу пятых суток диаметр бластодиска достигал 2,2—2,5 мм, а длина закладывающегося зародыша 0,8—1,1 мм. Чувствительность икры в начале гаструляции возрастает и в это время ее необходимо оберегать от каких бы то ни было толчков. 5 этап. Формирование головы и туловища зародыша. На ис
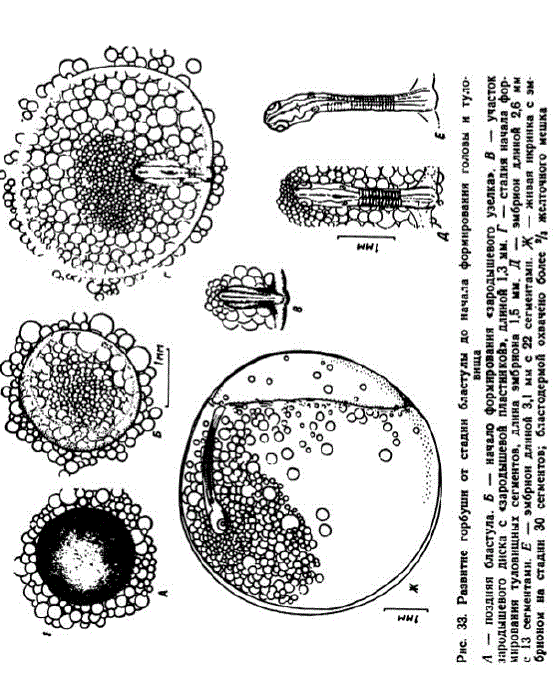
ходе 10-х сут. (82-86 градусо-дней) у эмбрионов образовались первые сомиты и начали формироваться мозговые пузыри. Диаметр зародышевого диска к этому времени увеличился до 3,5— 4 мм, эмбрионы достигли длины 1,5 мм или немного большей (рис. 33, Г). В условиях Лесного завода первые сомиты появлялись в конце 6-х сут.
На Соколовском заводе в 12 сут. (113,6 градусо-дней) эмбрионы имели длину 2,4—2,7 мм. У более мелких из них насчитывалось 5—7 сомитов. К этому времени четко обозначились зачатки глазных бокалов. В хвостовом конце можно было видеть купферов пузырек. Бластодерма охватывала участок поверхности желточного мешка диаметром 4,5—4,7 мм. У зародышей с 13 сомитами в глазах четко выявлялись хрусталики, заложились слуховые пузырьки, днфференцировались зачаток головы и туловища
(рис. 33, Д).
Форд и Ньюстед (Ford, Newstead, 1958) отметили возникновение нефротома и пронефрических складок у эмбрионов длиной 3,33 мм, имевших 18 сомитов. Нефротом располагался в области 6—7-го сомитов. В это же время под хордой обнаруживалось продольное плотное скопление клеточной массы, представляющее собой зачаток основных кровеносных сосудов.
В нашей серии эмбрионы длиной 3 мм имели 22 сомита (рис. 33, E). Продолжали дифференцироваться отделы головного мозга, наметились энцефаломеры. Слуховые пузырьки в это время отстояли далеко от глаз, головной отдел оставался большим.
В возрасте 13 сут. (122,7 градусо-дня) эмбрионы достигали 3,5—4 мм. У них насчитывалось до 33—35 сомитов (рис. 33, Ж). Начала формироваться хвостовая почка (несколько раньше, чем у кеты). Под передним отделом головы стала образовываться перикардиальная полость. Разрастающаяся бластодерма закрыла больше половины желточного мешка. С этого времени чувстви
6 этап. Обособление задней части туловища от поверхности желточного мешка. При развитии икры в Соколовском инкубаторе новый этап начался в возрасте около 17 сут. (155—160 градусо-дней), а в Лесном — вначале 13-х сут. Эмбрионы имели длину 42—4,5 мм.
После того как сформировался задний отдел кишки, анус и начали появляться сегменты хвостового отдела, задний участок туловищного отдела стал постепенно отделяться от желточного мешка. Этому процессу сопутствовало развитие преанальной плавниковой складки. У горбушн анус образовывался на уровне 40—43 сегментов. В начале этапа появлялись зачатки жаберных крышек, обозначились обонятельные плакоды. При длине эмбрионов 4,76 мм отмечено начало гистологической дифференцировки элементов пронефроса. У эмбрионов длиной 5,24 мм обнаруживались пронефрические клубочки, а максимального развития пронефрос достигал при длине 16,66 мм (Ford, Newstead, 1958).
В возрасте 17 сут. у отдельных эмбрионов были замечены слабые сокращения миотомов и сердечной трубки. У таких зародышей еще не завершилась эпиболия и сохранялась небольшая овальная желточная пробка. В условиях же Лесного завода эмбрионы приобретали подвижность после завершения эпиболии. Возможно, в первом случае развитие подвижности стимулировалось более низким содержанием в воде кислорода.
Перед закрытием желточной пробки чувствительность икры к механическим воздействиям резко возрастала.
7 этап. Развитие подкишечно-желточной системы кровообращения. Появление тока крови отмечалось в конце 19-х — середине 20-х сут. развития (178—183 градусо-дня). Такие эмбрионы имели длину 5,5—6,0 мм. В возрасте 19 сут. у них насчитывалось до 58, а в 19,5 сут. — 61—62 миотома (рис. 34, A). В условиях Лесного
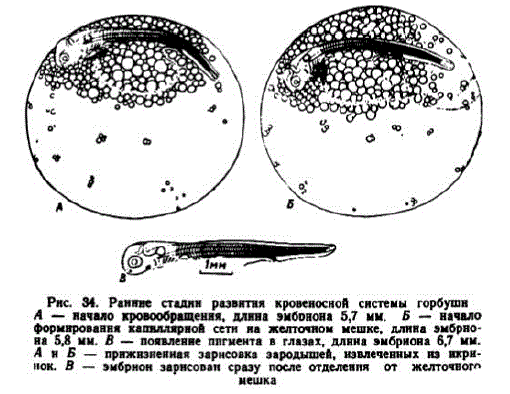
Рис. 34. Раннне стадин развития кровеносной системы горбушн A - начало кровообращення, длина эмбонона 5,7 мм. Б — начало формнровання капиллярной сети на желточном мешке, длнна эмбрнона 5,8 мм. В — появление ппгмента в глазах, длнна эмбриона 6,7 мм.
н Б — прижнзненная зарнсовка зародышей, извлеченных из икринок. B — эмбрион зарнсован сразу после отделения от желточного LIELILIKE
завода кровообращение начиналось в 14—15-суточном возрасте. В это время температура воды к утру стала снижаться до 10".
Кровообращение у горбуши начиналось на 1—2 сут. позже, чем у симы, инкубировавшейся в том же аппарате Соколовского инкубатора. Голова у таких эмбрионов начала отделяться от желточного мешка. Слуховые пузырьки отстояли от глаз на рас" стоянии, равном примерно ширине 9 туловищных миотомов. Значительно увеличился объем головного мозга. Наметилась ротовая Воронка. Появились зачатки жаберных крышек. Начали формиро
143
ваться обонятельные ямки. У основания первых трех миотомов, немного в стороне от туловища, появились зачатки грудных плавничков. Рассматривая под микроскопом живых зародышей в проходящем свете, чуть выше и позади грудных плавничков можно Gыло видеть первые клубочки пронефроса. Формируется непарная плавниковая складка, лучше выраженная снизу. Преанальная складка тянется вдоль 16—17 миотомов.
Кровь на этой стадии имела светлую окраску. Поток крови по спинной аорте достигал первых хвостовых сегментов, направлялся книзу, огибал заднюю кишку и по подкишечной вене выходил на левую сторону желточного мешка. Здесь подкишечно-желточная вена описывала неболыпую дугу и несла кровь к сердцу. У горбуши кровообращение может начинаться и на несколько более ранней стадии развития. На том же Соколовском заводе в 1953 г. движение крови впервые было обнаружено в возрасте 16-17 сут. (150—160 градусо-дней). Такие эмбрионы имели длину 5,4—5,6 мм и 58—60 миотомов, из которых 42—43 туловищных. У некоторых эмбрионов этой серии движение крови начиналось перед завершением эпиболии. Ускорение функционирования кровеносной системы мы связываем с несколько худшими условиями аэрацин икры.
В дальнейшем кровью снабжается все больший участок хвостового отдела. Подкишечно-желточная вена опускается ниже и на желточном мешке появляются тонкие капилляры (рис. 34, Б). Эмбрионы поводят хвостом то в одну, то в другую сторону. В условиях Соколовского завода этап длился 2,5—3 сут., несколько дольше, нежели у симы. После завершения эпиболии икринки приобретают высокую стойкость к толчками вибрации.
8 этап. Возникновение кардинальных вен и смешанного подкишечно-желточного и печеночно-желточного кровообращения. К исходу 22-х и в пачале 23-х сут. (206—211 градусо-дней) длина эмбрионов достигла 6,4—6,6 мм, число миотомов увеличилось до 67—68, из которых 23—24 располагались позади ануса. Количество эритроцитов увеличилось и кровь приобрела яркую красную окраску. Кровью в начале этапа снабжались 10—12 хвостовых сегментов. Из хвостовой вены кровь стала поступать не только в подкишечную вену, но и в образовавшуюся заднюю кардинальную вену. На желточном мешке позади туловища эмбриона появилось более 10 извитых капилляров, между которыми имелись анастомозы. Подкишечно-желточная вена опустилась, стала длиннес, собирает кровь из сосудовлевой и правой сторон желточного мешка и несет ее к сердцу.
Появление меланина в глазах у эмбрионов разных серий отмечалось в возрасте 22 и 23 сут. (213 и 215 градусо-дней), когда они имели длину 6,5—6,9 мм и 73—75 миотомов (рис. 34, B). Сообщение о начале пигментации глаз при 330 градусо-днях и длине эмбрионов 9,72 мм (Иевлева, 1951) ошибочно.
На описываемой стадии кончик хорды немного изогнулся
144
кверху. Непарная плавниковая складка увеличилась. Заметно подросли грудные плавнички и их основания приняли наклонное положсние. В статоакустических органах начали формироваться полукружные каналы; расстояние между ними и глазами сократилось. Голова стала заметно распрямляться.
Кровью снабжаются две пары жабер. Образовались капилляры на верхней поверхности головы. Хвостовая артерия приблизилась к последним сегментам. Появились первые сегментальные сосуды. Из печени на левую и правую стороны желточного мешка выходят печеночные вены, по которым кровь течет к сосудам, образованным подкишечно-желточной веной. Сеть капилляров, снабжаемых печеночными венами, быстро разрастается, тогда как кровоснабжение желточного мешка через подкишечную вену все более ослабевает. С увеличением эмбрионов и развитием сети капиллярных сосудов быстро возрастает потребление кислорода. Если тысяча 26-суточных зародышей на стадии слабой пигментации глаз при температуре 62-69 за час потребляли 0,3 мг кислорода, то через неделю, при 7,9—8,3", эта величина возросла no 0.6 Mr (Wickett, 1954).
9 этап. Формирование печеночно-желточной системы кровообращения. Подкишечно-желточная вена запустевала на 2829-е сут. (264—273 градусо-дня), когда эмбрионы достигали длины 7,5—8,2 мм. С этого времени желточный мешок снабжается кровью только печеночными венами. У таких эмбрионов заметно удлинился хвостовой отдел. Число миотомов в нем уменьшилось на 2—3. На спине непарная плавниковая складка разрослась до первых сегментов и заметно расширилась. Основания грудных плавников приняли полувертикальное положение. Увеличилась высота головы. Глаза стали серыми. В слуховых пузырьках продолжалось формирование полукружных каналов; многочисленные мелкие отолиты объединились в крупные.
В начале этапа кровью снабжались 3 пары жабер. Развились сосуды затылочной области. Сегментальные сосуды появились почти во всех туловищных миотомах. Почти вся поверхность желточного мешка, за исключением небольшого участка ниже головы, была покрыта капиллярами. С его правой стороны кровь стала направляться к сердцу по правой ветви краевой желточной вены. Задний отдел кишки был оплетен капиллярами задней брыжеечной артерии, из которых кровь поступала в подкишечную вену. Представление об уровне дифференцировки эмбрионов в конце этапа дает рис. 35, A (31 сут., 293 градусо-дня). Такие эмбрионы время от времени наклоняют тело то на левый, то на правый бок и из стороны в сторону плавно поводят хвостом.
10 этап. Дифференцировка верхних и нижних конусов миотомов. Верхние и нижние конусы миотомов в хвосте большинства эмбрионов появились в возрасте 32 сут. (303-градусо-дня) при длине 10,6—11,5 мм. У некоторых зародышей они были замечены На сутки раньше, у других позже. На Лесном заводе 10-й этап
начинался на 29—30-е сутки. Такие эмбрионы имели крупные черные, иридирующие глаза. Жаберные крышки прикрыли перв жабру. В обонятельных отверстиях наметились перемычки. На месте будущего анального плавника появились мускульные почки. Слабым расширением плавниковой складки и скоплением мезенхимной ткани обозначилось место закладки спинного плавника. Основания грудных плавников приобрели почти вертикальное положение и плавнички стали снабжаться кровью. Функционировали четыре пары жаберных артерий. Сегментальные сосуды развились во всех хвостовых миотомах. Краевая вена опустилась на вентральную сторону желточного мешка. В просвете кишки появился желто-зеленоватый пигмент.
На 34-е сутки (323 градусо-дня) зачатки верхних и нижних конусов имелись на всех миотомах (рис. 35, Б). С этого времени у извлеченных из оболочки эмбрионов желточный мешок принимает яйцевидную форму. Капиллярами покрылся весь желточный мешок. Длина капилляров на 1 мм2 его поверхности достигала 3,5 мм. Замечено ускорение слияния мелких и образование круп
В 38 сут. (370 градусо-дней) эмбрионы имели длину 12,6— 13,3 мм. Их грудные плавники приобрели способность двигаться. Начался процесс формирования гипуралий. В основании лопасти хвостового плавника образовались первые петли капилляров. Капилляры появились также вдоль миосепт первых туловищных M HOTOMOB,
К 40 сут. (392 градусо-дня) эмбрионы выросли до 13,5— I4,5 мм. Головы зародышей стали удлиняться. Нижняя челюсть достигла вертикали, опущенной от середины глаз. Жаберные крышки прикрыли две пары жабер. Начали функционировать гиоидные сосуды. На верхней поверхности головы развилась густая сеть капилляров. Протяженность капилляров на 1 мм? поверхности желточного мешка возросла до 5 мм. Увеличился объем мочевого пузыря. У живых эмбрионов под микроскопом можно было видеть появившиеся железки вылупления. В спинном плавнике четко обозначились мускульные почки. В хвостовом плавнике появились 3—4 лепидотрихии и увеличилась дорзальная часть лопасти. В течение этапа наблюдался быстрый рост хвостоВого отдела, в котором шло формирование позвонков. Продолжала расширяться непарная плавниковая складка. Заложились брюшные плавники. На поверхности головы обнаруживались первые меланофоры (рис. 35, У горбуши хроматофоры появляются позже, чем у всех других тихоокеанских лососей.
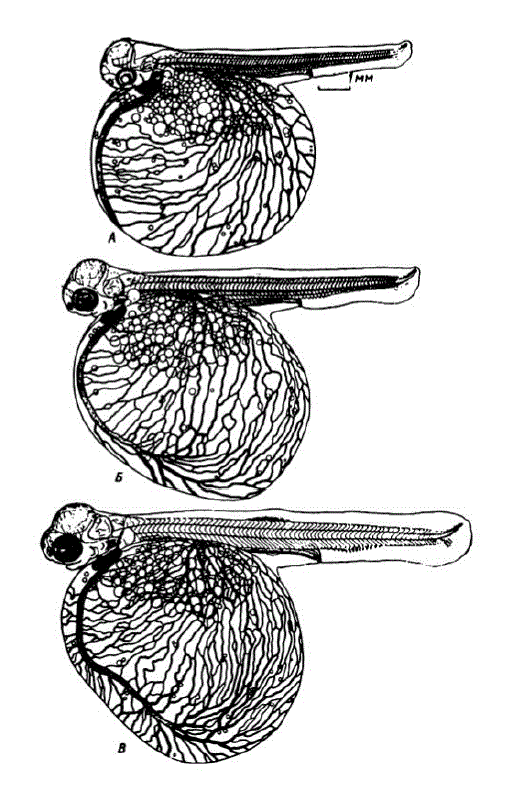
Рис. 35. Горбуша на различных стадиях формировання желточного вообращения нразвнтия верхних н нижних конусов миотомов - возраст 31 сут. (293 градусо-дня), длина эмбриона 104 мм. возраст 34 сут. (323 градусо-дия), длина эмбриона 11,4 мм. - возраст 40 сут. (392 градусо-дня), длина эмбриона 14,1 мм
У эмбрионов длиной 14,7 мм завершается формирование хрящевого основания черепа, образуются его боковые стенки, наблюдается охрящевение жаберных дуг и зачатков пояса грудных плавников (Иевлева, 1951).
В течение этапа под оболочкой эмбрионы двигаются мало, изредка делают по нескольку взмахов грудными плавниками. Время от времени они возбуждаются и совершают серию волнообразных движений. В конце этапа некоторые особи начинают слабо двигать челюстями.
11 этап. Развитие подвижности челюстей, жаберных крышек, завершенненнкубации. В начале этапа висцеральный аппарат приобретает подвижность. У значительной части эмбрионов это происходило в возрасте 45—47 сут. (445—466 градусо-дней), когда "они достигали длины 15 мм и более. К этому возрасту слуховые капсулы максимально сблизились с глазами. Жаберные крышки закрыли три пары жабер, на которых появились зачатки лепестков. Разрастающаяся нижняя челюсть почти достигла линии переднего края глаз (рис. 36, A).
Увеличившийся хвостовой отдел составляет около 35—36% всей длины. Началось формированне опорных элементов спинного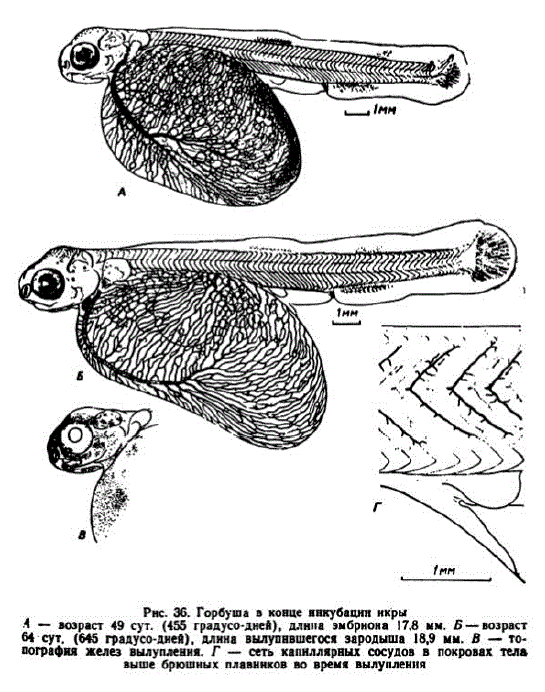 L жирового плавника. Лопасти грудных плавников стали крупными, а их основания сузились. Идет окостенение элементов плечевого пояса. На спине эмбриональная плавниковая складка стала укорачиваться.
L жирового плавника. Лопасти грудных плавников стали крупными, а их основания сузились. Идет окостенение элементов плечевого пояса. На спине эмбриональная плавниковая складка стала укорачиваться.
Продолжает возрастать густота сосудистой сети желточного мешка, увеличивается сеть капилляров на голове, в покровах тела, хвостовой лопасти. Межсегментальные капилляры развились по всему туловищу и на первых хвостовых миотомах. В покровах головы меланофоров стало больше, они потемнели, появились мелкие оранжево-красные липофоры.
В возрасте 56 сут. (560 градусо-дней) эмбрионы достигали длины 18 мм и более. Они приобрели способность двигать глазами. Жаберные крышки разрослись до последней пары жабер. Установилась связь ротовой и жаберной полостей. Жаберные лепестки удлинились до 0,15—0,17 мм, в них, так же как в псевдобранхиях, появились капилляры. Хвостовой отдел увеличился до 36,5—37,2% длины зародышей. В хвосте появилось 13—14 лепидотрихий. Наметился контур жирового плавничка. В это время хорошо видны железки вылупления, которых у горбуши особенно много (рис. 36, B).
Ко времени вылупления капиллярная сеть желточного мешка достигает максимального развития. На 1 мм2 его поверхности капилляры имеют длину до 6,5 мм; это меньше, чем у родственных видов. Сосуды, отходящие от сегментальных артерий, образуют довольно густую сеть вдоль спинного мозга и вблизи поверхности тела (рис. 36, Г). Меланофорами покрылась верхняя поверхность головы, затылочная область; на спине они мельче,
светлее, располагаются редко. Красноватые мелкие липофоры обильнее также на голове, на спинке редки, позади спинного плавника они появились в основании плавниковой складки, причем в несколько большем количестве на месте формирующегося жирового плавника.
При 10,4—10,7° этап длился 20—23 сут.
12 этап. Пассивное состояние свободных зародышей. Первые зародыши вылупились на 64-е сут.; через 2 дня вылупление стало массовыми закончилось на 70-е сут. (от 645 до 708 градусо-дней); "ಲ್ಗಙ್ಗ за время инкубации менялась в пределах 10,7— 8,4". В одинаковых условиях икра горбуши инкубировалась дольше, чем икракеты (по числу суток примерно на 20%, по количеству градусо-днейна 12%).
Как и в реках, где в зависимости от времени нереста и температуры воды икра инкубируется от 1—1,5 до 4—5 месяцев, на рыбоводных заводах эти сроки также подвергнуты значительным колебаниям. На сахалинских заводах массовое вылупление происходит в возрасте от 50 до 150 сут., а число градусо-дней колеблется в пределах 300—740. Изменение сроков инкубации икры в зависимости от времени ее сбора в условиях Лесного завода, снабжаемого речной водой, приведены в табл. 10. Икра осенних сборов развивается при низкой температуре очень долго, однако число градусо-дней оказывается меньшим; сильно растягиваются и сроки вылупления.
Таблица 10 продолжительность инкувлции икры Горвуши PA3HX CPOKOB OESOPA HA JEOTHOM PbN6OBOHOM
При вылуплении зародыши описываемой серии имели длину от 18,2 до 21,7 мм (размер абсолютного большинства из них составлял 19—21 мм; измерялись живые объекты). Зародыши весили 119,5—174,0 мг (М=143,2+17,1), причем вес желтка составлял от 74 до 84,3% общего веса. В одной серии мелкой икры зародыши вылуплялись при длине от 16 до 18,5 мм (17,49+ #? B HKpHHках диаметром около 6,7 мм ко времени вылупления эмбрионы достигали длины 18,3—19,9 мм, а в икринках диаметром 7,4 мм— 20.4—21,7 мм (измерения прижизненные).
Вылупившиеся зародыши имеют массивную голову с крупными подвижными глазами. Их жабры с хорошо развитыми лепестками и зачатками первых жаберных тычинок закрывают жаберные крышки. Имеются лучи жаберных перепонок. Четко выражены верхнечелюстные кости. В обонятельных отверстиях завершается формирование перемычек. Желточный мешок после вылупления удлиняется, его задний конец простирается до анального отверстия и немного опускается (рис. 36, Б). Ко времени вылупления основная массажировых капель сливается в одну крупную, диаметр которой достигает 2,5 мм и более. Верхнюю часть желточного мешка начинают покрывать разрастающиеся миотомы.
У вылупившихся зародышей насчитывалось 66—68 миотомов, из которых в хвосте — 25—27. Постанальнос расстояние составляло около 36—37% всей длины. В хвосте образовалось 20—22 костных луча, средние из которых имели по 3 и даже 4 членика. При раннем вылуплении в хвосте бывает менее 18 лучей и в них меньше члеников. К концу инкубации непарная плавниковая складка заметно редуцируется, благодаря чему более четко выявляются контуры непарных плавников. В них хорошо обозначаются зачатки лепидотрихий и птеригиофор. Преанальная плавниковая складка, наоборот, продолжает расти. Грудные плавнички стали крупными, заметно удлинились и зачатки брюшных плавников. В момент вылупления по Y888 развития органов боковой линии горбуша сходна с кетой. Пигментирована она слабее других ви
LOEB,
Желудочно-кишечный тракт во время вылупления представляет собой почти прямую трубку, в которой некоторое утолщение стенок и наметившийся изгиб обозначают место формирования желудочного отдела. Зачаток плавательного пузыря представлен пальцеобразным выростом, длиной немного более 1 мм.
После вылупления зародыши успокаиваются. По уровню дифференцировки рецепторных органов, отношению к внешним воздействиям, поведению они не отличаются от других лососей. Вышедшие из икры зародыши при температуре 8—9° в относительно пассивном состоянии находятся около недели. При температуре 3—4° этот срок почти удваивается.
13 этап. Формирование непарных, брюшных плавников и плавательного пузыря. В начале этапа свободные зародыши приобретают положительную реакцию на течение и прикосновение к посторонним предметам, отрицательную реакцию на свет. В питомниках с ровным дном, как это нами наблюдалось на Адотымовском заводе, они стремятся переместиться к местам поступления воды и образуют здесь, а также в углублениях дна скопления. В питомниках с галечным покрытием н умеренным течением зародыши распределяются равномернее. От яркого освещения, стука, механического раздражения свободные зародыши возбуждаются и могут подняться в толщу воды, но, отягощенные огромным желточным мешком, удерживаются на плаву недолго и вскоре
опускаются на дно. Следует заметить, что при низкой температуре изменение поведения проявляется нечетко.
К началу этапа рот принимает полуконечное положение (рис. 37, A). Непарная плавниковая складка впереди спинного плавника резорбируется; преанальная складка по-прежнему остается крупной. В хвосте начинают ускоренно расти верхние и нижние лучи, благодаря чему в его лопасти образуется выемка. Формируются лучив грудных и брюшных плавниках.
После вылупления убыстряется резорбция желтка. В течение этапа увеличивается число жаберных тычинок, их размер, растут разветвленные жаберные лепестки. Дифференцируются отделы желудочно-кишечного тракта, закладываются пилорические придатки. Быстро растет зачаток плавательного пузыря. Меланофоры продолжают оставаться мелкими, расположены редко и только на дорзальной стороне тела. В конце этапа в коже, кроме имевшихся оранжево-красных липофоров, обнаруживаются более мелкие хроматофоры лимонно-желтого цвета. Меланофорами интенсивно пигментируется брюшина, они покрывают головной и спинной мозг, располагаются вдоль основных кровеносных сосудов.
Горбушу отличает раннее появление зародышевых клеток и быстрое развитие гонад, которые первоначально у всех особей формируются как яичники (Robertson, 1953; Персов, 1965, 1966, 1968). По данным Г. М. Персова, через 7—10 дней после вылупления в гонадах обнаруживаются овоциты периода протоплазматического роста. В разных температурных условиях передифференцировка пола и формирование семенников происходит в период от 30 до 123 дней после вылупления (582—906 градусо-дней), при длине личинок от 24 до 30 мм.
Смотрите также
Бассейновый метод выращивания лососевых рыб